Эндотипы и фенотипы астмы — от алгоритма обследования до подбора терапии
Информация только для специалистов в сфере медицины, фармации и здравоохранения!
С 18 по 20 февраля 2015 г. в Москве прошел XIII Международный конгресс Российской ассоциации аллергологов и клинических иммунологов, в рамках которого состоялся симпозиум «Эндотипы и фенотипы астмы — от алгоритма обследования до подбора терапии», посвященный современным подходам к диагностике и лечению пациентов с бронхиальной астмой.
С вступительной речью выступил председатель симпозиума д.м.н., профессор, заведующий кафедрой аллергологии и пульмонологии Иркутской государственной медицинской академии последипломного образования, главный внештатный аллерголог-иммунолог Сибирского и Дальневосточного федерального округа Борис Анатольевич Черняк:
Последние несколько десятилетий мы имеем дело с персонализированными подходами в медицине. GINA в последнюю свою редакцию внесла небольшое, на первый взгляд, но очень существенное изменение в определение бронхиальной астмы (БА).

Wenzel предложила на основании молекулярного эндотипирования в рамках эндотипов Th3 и не Th3 воспаления выделять определенные фенотипы БА. Для ТН2 это аллергическая астма (в т. ч. ее вариант – атопическая астма), аспириновая БА, которая может быть самостоятельным аллергическим заболеванием или сочетаться с атопией. Сюда же частично относится синдром перекреста (атопическая БА в сочетании с ХОБЛ). Это все — ТН2 эндотип, который объединяет различные варианты бронхиальной астмы и эозинофильного воспаления. Не ТН2 эндотип — это астма с очень поздним дебютом, астма с ожирением, астма у курильщиков или астма нейтрофильная у некурящих людей.
Почему же нам так важны эти фенотипы и эндотипы бронхиальной астмы? С моей точки зрения, ответ очевиден: разные этиологические факторы и факторы риска, разное течение заболевания обуславливают разные подходы к терапии и разный ответ на лечение и, соответственно, прогноз.
Д.м.н., главный аллерголог Министерства здравоохранения Республики Татарстан, заведующий кафедрой аллергологии Казанской государственной медицинской академии, профессор Рустем Салахович Фассахов выступил с докладом «Атопическая астма:
клиническая характеристика и возможности современной терапии»:
Атопия — это генетическая предрасположенность к аллергическим заболеваниям, обусловленная гиперпродукцией IgE-антител к аллергенам. Важнейшим отличительным признаком атопии является наследственная предрасположенность. Хорошо знакомые причины атопической астмы — это домашние аллергены, источниками которых служат животные, клещи домашней пыли, споры плесневых грибов и т. д.
д.
Какие клинические особенности выделяются у атопической БА? Считается, что типичен старт заболевания в детском возрасте, но на самом деле так происходит не всегда. В большинстве случаев воспалительный процесс в бронхах носит эозинофильный характер. Началу заболевания, как правило, предшествует аллергический ринит. Он выявляется практически у 100% больных с атопической БА.
Обсудим GINA в редакции 2014 г. Документ можно назвать революционным. Он категорически отличается от своих предыдущих версий. Надо сказать, что главное отличительный признак новой редакции — это приближенность к жизни. Большинство наших врачей довольно давно используют на практике рекомендации, впервые появившиеся в 2014 г.
На рисунке 1 вы можете увидеть все основные изменения, появившиеся в GINA 2014 г. Как уже было сказано ранее, наконец-то в определении астмы прозвучало, что это гетерогенное заболевание. Среди основных изменений в GINA – так же акцент на нефармакологическое влияние на БА. Если говорить о диагностике астмы, отмечено, что симптомы заболевания могут появляться при контакте с аллергенами. Для атопической астмы связь заболевания с контактом с аллергенами — очень важный признак.
Если говорить о диагностике астмы, отмечено, что симптомы заболевания могут появляться при контакте с аллергенами. Для атопической астмы связь заболевания с контактом с аллергенами — очень важный признак.
Один из моментов диагностики БА — наличие вариабельной обструкции. Для этого мы исследуем функцию внешнего дыхания пациента. Если первоначальное исследование дало отрицательный результат, повторите его в период наличия симптомов (сезон цветения, контакт с животными), либо после отмены бронхолитических препаратов.
Контроль астмы — это две составляющие: контроль факторов риска заболевания и учитывание проблем, связанных с проведением терапии. Факторы риска: риносинусит, ГЭРБ, ожирение, обструктивное апноэ во сне, депрессия. Все это может утяжелять симптомы БА и ухудшать качество жизни. Что подразумевается под проблемами, связанными с проведением терапии? Приверженность к терапии, правильность выполнения техники ингаляций, отношение больного к своему заболеванию и т.
Как оценивается степень тяжести астмы? Ретроспективно по объему проводимой для контроля симптомов и обострений терапии в течение нескольких месяцев. Тяжесть БА может изменяться с течением времени. Например, терапия ринита существенно улучшает состояние больного БА. Что важно, когда мы начинаем оценивать проводимую терапию? В зависимости от результатов лечения проводится коррекция терапии, в т. ч. нефармакологическая. То есть соблюдение элиминационного режима, техника проведения ингаляций и т. д.
Что нового в вопросах терапии БА в последней редакции GINA? На второй ступени появляются группы препаратов (антагонисты лейкотриеновых рецепторов и КДБА), которые могут быть использованы у больных с БА и аллергическим ринитом. Периодический прием ИГКС и КДБА по потребности у больных с чисто пыльцевой БА при отсутствии симптомов вне сезона; назначение ИГКС сразу с появлением симптомов и продолжение терапии не менее 4 недель после завершения сезона. Как я уже сказал, это — новые аспекты терапии БА в редакции GINA 2014, но вполне привычные для наших специалистов.
Как я уже сказал, это — новые аспекты терапии БА в редакции GINA 2014, но вполне привычные для наших специалистов.
На третьей ступени есть серьезные изменения, касающиеся базовой терапии. Например, комбинация низких доз ИГКС плюс формотерол — как препараты поддержки, так и по потребности.
Астма — это заболевание вариабельное. Обострения могут возникать даже у пациентов с контролируемой БА под воздействием тех или иных триггеров. Но обострения можно предотвратить, увеличив дозу ИГКС в 4 раза. То есть причина увеличения дозы ИГКС — массивный и длительный контакт с триггером (например, в сезон цветения). Причина для увеличения дозы ДДБА — превентивное использование перед краткосрочными контактами с аллергенами.
На 4 ступени на первый план выходит терапия комбинированными препаратами ИГКС с ДДБА. Одна из таких фиксированных комбинаций — это Форадил Комби, содержащий в себе формотерол (12 мг) и будесонид (200) мг. Он прекрасно вписывается в рекомендации GINA 2014. Посмотрите на рисунок 2, где отражено место Форадила Комби в лечении астмы. Его доза отлично встраивается и в терапию третьей ступени, и четвертой, и пятой. На пятой ступени мы можем добавлять этот препарат по потребности, когда в этом есть необходимость. Дозу Форадила Комби можно как повышать, так и понижать. Раздельные ингаляции позволяют нам изменять дозировку каждого компонента в отдельности. Форадил Комби отлично подходил как для терапии впервые выявленной БА, так и в том случае, когда мы вынуждены больному менять один препарат на другой в силу тех или иных обстоятельств.
Посмотрите на рисунок 2, где отражено место Форадила Комби в лечении астмы. Его доза отлично встраивается и в терапию третьей ступени, и четвертой, и пятой. На пятой ступени мы можем добавлять этот препарат по потребности, когда в этом есть необходимость. Дозу Форадила Комби можно как повышать, так и понижать. Раздельные ингаляции позволяют нам изменять дозировку каждого компонента в отдельности. Форадил Комби отлично подходил как для терапии впервые выявленной БА, так и в том случае, когда мы вынуждены больному менять один препарат на другой в силу тех или иных обстоятельств.
Пятая ступень терапии БА — очень важная. Первая рекомендация — направление больного к специалисту на обследование. О чем это говорит? Первые четыре ступени лечит врач первичного клинического звена. И только на 5 ступени вы отправляете больного к пульмонологу или аллергологу. Опять же эксперты напоминают нам, что необходимо проверить технику использования ингалятора.
На 5 ступени возможно использование препарата омализумаб (антагонист лейкотриеновых рецепторов) у пациентов с подтвержденной аллергической БА, не поддающейся контролю терапией четвертой ступени. Он назначается тем больным, у которых не достигается контроля над заболеванием даже при самой адекватной, современной, правильно выполняемой терапии, в т. ч. комбинированным ингаляционным препаратом. Исследования INNOVATE и PERSIST показали лучший контроль астмы, снижение количества обострений и улучшение качества жизни у получающих препарат больных с аллергической астмой.
Он назначается тем больным, у которых не достигается контроля над заболеванием даже при самой адекватной, современной, правильно выполняемой терапии, в т. ч. комбинированным ингаляционным препаратом. Исследования INNOVATE и PERSIST показали лучший контроль астмы, снижение количества обострений и улучшение качества жизни у получающих препарат больных с аллергической астмой.
Резюмируя вышесказанное, хочу еще раз подчеркнуть, что большинство из новых рекомендаций GINA мы уже давно успешно используем. И это говорит о том, что мы ведем работу в правильном направлении, а экспертные рекомендации максимально приближаются к существующим реалиям.
К.м.н., доцент кафедры аллергологии и пульмонологии Иркутской государственной медицинской академии последипломного образования Ирина Ивановна Воржева выступила с докладом «Аспириновая астма: алгоритм диагностики и особенности лечения»:
Аспириновая бронхиальная астма (БА) была описана еще в начале прошлого века. Во второй половине ХХ столетия она получила название аспириновой или астматической триады, что отражало три главных признака этой болезни: наличие астмы, риносинусита (чаще всего полипозного) и непереносимости ацетилсалициловой кислоты (АСК), которая проявляется приступами удушья и назальной симптоматикой. В начале ХХI века появился другой термин: аспирин-индуцированное респираторное заболевание. Этот термин отражает сложившееся понимание, что астма и риносинусит сочетаются не случайно, а имеют очень тесные патогенетические связи, другими словами: одни воздушные пути, одна болезнь. При этом описано уже достаточное количество случаев развития непереносимости АСК у пациентов с риносинуситом без астмы. И, наконец, возникает относительно новое понятие — НПВП-индуцированное респираторное заболевание или NERD (от англ. nonsteroidal anti-inflammatory drugs-exacerbated respiratory disease), которое подчеркивает, что триггером может являться не только АСК, но и НПВП, относящиеся к ингибиторам циклооксигеназы 1 типа (ЦОГ-1).
Во второй половине ХХ столетия она получила название аспириновой или астматической триады, что отражало три главных признака этой болезни: наличие астмы, риносинусита (чаще всего полипозного) и непереносимости ацетилсалициловой кислоты (АСК), которая проявляется приступами удушья и назальной симптоматикой. В начале ХХI века появился другой термин: аспирин-индуцированное респираторное заболевание. Этот термин отражает сложившееся понимание, что астма и риносинусит сочетаются не случайно, а имеют очень тесные патогенетические связи, другими словами: одни воздушные пути, одна болезнь. При этом описано уже достаточное количество случаев развития непереносимости АСК у пациентов с риносинуситом без астмы. И, наконец, возникает относительно новое понятие — НПВП-индуцированное респираторное заболевание или NERD (от англ. nonsteroidal anti-inflammatory drugs-exacerbated respiratory disease), которое подчеркивает, что триггером может являться не только АСК, но и НПВП, относящиеся к ингибиторам циклооксигеназы 1 типа (ЦОГ-1). И поэтому при NERD наблюдается очень широкий перекрест реакции гиперчувствительности с различными неселективными НПВП.
И поэтому при NERD наблюдается очень широкий перекрест реакции гиперчувствительности с различными неселективными НПВП.
В настоящее время выделяют пять вариантов чувствительности к АСК и НВПВ. NERD относится к неаллергической, неиммунологической гиперчувствительности, в основе которой лежат генетически обусловленные нарушения метаболизма арахидоновой кислоты.
НПВП — лишь часть триггеров аспириновой БА. Кроме них обсуждаются и другие различные пусковые механизмы: персистенция респираторных вирусов, стимуляция антигенами золотистого стафилококка и т. д. Распространенность NERD в общей популяции невелика, а вот от общего количества больных БА по данным анамнеза NERD составляет 11%, в то время как при проведении провокационных тестов частота его возрастает до 21—28%.
Типичная клиническая картина NERD включает БА, гиперпластический риносинусит, полипоз полости носа и придаточных пазух (это, правда, не является облигатным признаком) и характерную эозинофильную инфильтрацию слизистой респираторного тракта. Маркером этой болезни является гиперчувствительность к АСК и НПВП с проявлением симптомов астмы и/или ринита.
Маркером этой болезни является гиперчувствительность к АСК и НПВП с проявлением симптомов астмы и/или ринита.
Типичным возрастом дебюта аспириновой астмы считается 20—40 лет. Аспириновая астма нередко бывает вариантом поздно возникшей БА. Для нее характерно персистирующее, недостаточно контролируемое течение с частыми обострениями, в т. ч. вирусоиндуцированными. При тяжелом течении может формироваться необратимая бронхиальная обструкции и стероидная зависимость, нередко в сочетании с атопией (до 70% случаев).
Диагностика NERD в рутинной практике достаточно проста: это анамнез БА, сочетающийся с выраженной патологией верхних дыхательных путей, а так же респираторные симптомы, обостряющиеся после применения АСК и/или НПВП. Используются рутинные лабораторные исследования: общий анализ крови, риноцитограмма, где выявляется эозинофилия. Риноскопия позволяет обнаружить гипертрофию слизистой полости носа или полипоз. Компьютерная томография носа и придаточных пазух выявляет гиперпластичность или полипозный риносинусит.
Предположение об аспириновой БА должно, конечно, возникать при всех случаях БА, сочетающейся с полипозным риносинуситом. В тоже время можно даже у больных с отсутствием полипозного риносинусита заподозрить формирование NERD, если течение БА становится неконтролируемым на фоне применения АСК и НПВП.
Диагностика гиперчувствительности к НПВП базируется в первую очередь на данных анамнеза. Провокационный тест (золотым стандартом является пероральный) применяется при наличии клинических признаков NERD, но отсутствии четких указаний в анамнезе на гиперчувствительность к аспирину и НПВП. Кожные пробы с АСК и НПВП не проводятся, т. к. механизм гиперчувствительности при NERD не является иммунологическим.
Лабораторная диагностика пока не получила широкого распространения, включая и тест активации базофилов, т. к. связана с недостаточной доказательной базой в отношении чувствительности, специфичности и воспроизводимости предлагаемых методов.
Принципы лечения NERD складываются из противовоспалительной терапии патологий верхних и нижних дыхательных путей, из исключения, а чаще ограничения триггеров, поддерживающих тканевое воспаление (АСК, НПВП, природные салицилаты), снижения чувствительности к аспирину и НПВП. В некоторых случаях имеются показания к десенситизации АСК, а так же новое направление — терапия биологическими препаратами.
В некоторых случаях имеются показания к десенситизации АСК, а так же новое направление — терапия биологическими препаратами.
Фармакотерапия аспириновой БА полностью соответствует принципам ступенчатого подхода к лечению астмы. И поскольку аспириновая астма — это чаще всего среднетяжелое, а в большей степени и тяжелое персистирующее течение, то лечению, как правило, соответствует 3—5 ступени по GINA. Это комбинации ингаляционных глюкокортикостероидов (ИГКС) и длительно действующих β2-агонистов (ДДБА) на всех ступенях. И если этого недостаточно, то прежде чем повышать дозы, усиливают терапию антилейкотриеновыми препаратами. На 5 ступени добавляют омализумаб и низкие дозы пероральных ГКС. Точки приложения действия противовоспалительных препаратов показаны на рисунке 3. Кортикостероиды прерывают всю цепочку метаболизма активации арахидоновой кислоты в самом начале. Ингибиторы 5-липоксигеназы в РФ не зарегистрированы, поэтому мы чаще всего пользуемся антагонистами цистеиниловых рецепторов лейкотриена. Это препараты, которые действуют в первую очередь в отношении рецепторов лейкотриена D4, в меньшей степени С4 и Е4. Антилейкотриеновые препараты (монтелукаст) применяются как для лечения астмы, так и для риносинусита. Однако они используются только в комбинации с кортикостероидами. Монотерапия антилейкотриеновыми препаратами не позволяет при NERD достичь полноценного контроля над сложными механизмами воспаления. В тоже время добавление монтелукаста к назальным кортикостероидам уменьшает частоту рецидивов полипоза носа после функциональной операции.
Это препараты, которые действуют в первую очередь в отношении рецепторов лейкотриена D4, в меньшей степени С4 и Е4. Антилейкотриеновые препараты (монтелукаст) применяются как для лечения астмы, так и для риносинусита. Однако они используются только в комбинации с кортикостероидами. Монотерапия антилейкотриеновыми препаратами не позволяет при NERD достичь полноценного контроля над сложными механизмами воспаления. В тоже время добавление монтелукаста к назальным кортикостероидам уменьшает частоту рецидивов полипоза носа после функциональной операции.
Новые направления в лечении NERD — целевая терапия. Используются моноклональные антитела к IgE (омализумаб). Рандомизированное двойное слепое плацебо-контролируемое исследование показало уменьшение выраженности симптомов риносинусита и астмы (блокада носа, ринорея, одышка), улучшение обоняния и качества жизни, уменьшение размера полипов (по данным компьютерной томографии). В другом – открытом исследовании, кроме улучшения клинико-функционального статуса было показано еще и повышение толерантности к АСК.
Что же касаемо моноклональных антител к IL-5 (меполизумаб), то по данным рандомизированного двойного слепого плацебо-контролируемого исследования отмечалось уменьшение симптомов полипозного риносинусита и размера самих полипов, а так же снижение эозинофилии в мокроте у больных.
На рисунке 4 представлены факторы риска обострений БА и развития фиксированной обструкции.
Следует сказать, что NERD является вариантом болезни, требующим особых подходов к диагностике и лечению. Далеко не всегда наблюдается типичная астматическая триада, что затрудняет диагностику. АСК и НПВП широко применяются в терапии многих заболеваний, поэтому гиподиагностика NERD способствует прогрессированию БА и может представлять угрозу для жизни пациента.
Элиминация АСК/НПВП и природных салицилатов не приводит к ремиссии, так как механизмы воспаления при аспириновой БА обусловлены влиянием многих факторов.
Для достижения контроля аспириновой БА необходима одновременная терапия воспаления верхних и нижних дыхательных путей, что в результате позволяет снизить суммарную суточную дозу кортикостероидов.
Контролируемые исследования показали, что доставка ИГКС и ДДБА в комбинированном ингаляторе так же эффективна, как и прием каждого препарата по отдельности (GINA 2014). Современные комбинированные препараты ИГКС/ДДБА улучшают клинико-функциональный статус и качество жизни большинства пациентов, в том числе с тяжелой аспириновой БА.
Результаты российских исследований показали быстрое достижение контролируемого течения БА с помощью комбинации формотерола и будесонида (препарат Форадил Комби) при БА средней степени тяжести и при тяжелой форме БА (рис. 5–6).
Оценка техники ингаляции и использование ингаляторов, которые обеспечивают надежную легочную депозицию, имеют важное значение в достижении контроля астмы (рис. 7).
При плохо контролируемой астме используется сначала бронходилататор, а потом через некоторое время кортикостероид, тогда вероятность попадания его в мелкие дыхательные пути увеличивается. Кроме того, очень удобно корректировать дозировку, изменяя ее от самых высоких доз ингаляционных стероидов до сочетания с ДДБА и постепенно снижая до низких доз монотерапии только одним будесонидом. Возможность использовать Форадил Комби таким образом послужила основанием называть его конструктором для лечения БА. При этом, конечно же, важно помнить, что для эффективного и безопасного применения Форадила Комби необходимо тщательное обучение пациента и постоянный контроль за правильным применением препарата.
Кроме того, очень удобно корректировать дозировку, изменяя ее от самых высоких доз ингаляционных стероидов до сочетания с ДДБА и постепенно снижая до низких доз монотерапии только одним будесонидом. Возможность использовать Форадил Комби таким образом послужила основанием называть его конструктором для лечения БА. При этом, конечно же, важно помнить, что для эффективного и безопасного применения Форадила Комби необходимо тщательное обучение пациента и постоянный контроль за правильным применением препарата.
Б.А. Черняк, доклад «Бронхиальная астма и хроническая обструктивная болезнь легких: синдром перекреста»:
Бронхиальная астма и хроническая обструктивная болезнь легких — сочетание этих двух заболеваний известно давно. Но проблема настолько значима, что два коллективных эксперта в области пульмонологии — GINA и GOLD — издали недавно документ, где рассматривается именно этот вопрос. Определение синдрома звучит так: синдром перекреста бронхиальной астмы и ХОБЛ (СПАХ) характеризуется постоянным ограничением воздушного потока и рядом симптомов, которые связаны с астмой, а также признаками, характерными для ХОБЛ. Соответственно СПАХ диагностируется при наличии клинических признаков, соответствующих как БА, так и ХОБЛ.
Соответственно СПАХ диагностируется при наличии клинических признаков, соответствующих как БА, так и ХОБЛ.
Это сочетание по данным разных авторов и в разных популяциях варьирует от 15 до 55%. Речь идет не просто о сочетании двух распространенных и тяжелых заболеваниях. Синдром взаимного отягощения приводит к более тяжелому течению заболевания, чем просто при БА или ХОБЛ:
• более частые обострения,
• более низкое качество жизни,
• более быстрое снижение функции легких,
• более высокая смертность,
• больший объем использования ресурсов здравоохранения.
Каковы клинические особенности синдрома перекреста? Обычно это пациент старше 40 лет (хотя в целом это категория больных моложе, чем при изолированной болезни легких) с клинико-функциональными параметрами ХОБЛ, у которых ранее диагностировалась БА. ХОБЛ при СПАХ развивается в более молодом возрасте, даже при меньшей продолжительности и интенсивности курения. Выделяют 2 фенотипа СПАХ: эозинофильный и нейтрофильный, которые различаются ответом на терапию ИГКС.
Синдромальный подход к диагнозу сочетания БА и ХОБЛ построен на том, что учитываются симптомы, характерные для астмы, и симптомы, характерные для ХОБЛ (рис. 8). Если имеются более чем три симптома, характерных для астмы, и 3 и более, характерных для ХОБЛ, то, как правило, этого достаточно, чтобы весьма обоснованно заподозрить сочетание этих заболеваний и синдром перекреста БА и ХОБЛ.
Когда речь идет о СПАХ, то образ больного может быть следующим: во-первых, возраст начала, как я уже сказал, обычно ≥40 лет, но может быть дебют и ранее. Во-вторых, характер респираторных симптомов, в т. ч. одышки при физической нагрузке: эти симптомы постоянные, но сочетаются с выраженной вариабельностью. Функция легких: ограничение воздушного потока не полностью обратимо, но опять же имеется вариабельность. Вне обострений наблюдается постоянное ограничение воздушного потока (ОФВ1/ФЖЕЛ <0,7). В анамнезе заболевания, в семейном анамнезе часто ранее диагностированная БА, аллергия и/или вредные воздействия. Заболевание протекает с типичным прогрессированием и имеет высокую потребность в лечении. При этом даже на фоне терапии симптомы уменьшаются не полностью. Обострения могут быть частыми, но уменьшаются на фоне лечения. Сопутствующие заболевания способствуют ухудшению течения СПАХ. Лучевая диагностика проводится так же, как и при ХОБЛ. При исследовании мокроты обнаруживаются эозинофилы и/или нейтрофилы.
Заболевание протекает с типичным прогрессированием и имеет высокую потребность в лечении. При этом даже на фоне терапии симптомы уменьшаются не полностью. Обострения могут быть частыми, но уменьшаются на фоне лечения. Сопутствующие заболевания способствуют ухудшению течения СПАХ. Лучевая диагностика проводится так же, как и при ХОБЛ. При исследовании мокроты обнаруживаются эозинофилы и/или нейтрофилы.
На что следует обращать внимание из дополнительной диагностики? Еще до недавнего времени показатель бронхиальной гиперреактивности позиционировался как своего рода «водораздел» между БА и ХОБЛ. Бронхиальная гиперреактивность считалась признаком БА и была нехарактерна для ХОБЛ. На самом деле в последние годы стало ясно, что этот показатель не имеет диагностического значения. Просто при БА показатель бронихиальной гиперреактивности выше. Бронхиальная гиперреактивность имеет место почти в 90% случаев при ХОБЛ!
Актуальным при СПАХ является компьютерно-томографическое исследование высокого разрешения. Оно позволяет подчеркнуть, что имеет место не просто гиперинфляция легких, которая может отмечаться у больных астмой, не просто повышенная воздушность, а признаки эмфизематозных изменений. На рисунке 9 представлены методы дополнительного обследования.
Оно позволяет подчеркнуть, что имеет место не просто гиперинфляция легких, которая может отмечаться у больных астмой, не просто повышенная воздушность, а признаки эмфизематозных изменений. На рисунке 9 представлены методы дополнительного обследования.
Диагноз СПАХ строится на обнаружении клинических признаков и БА, и ХОБЛ, которые мы подтверждаем необратимостью бронхиальной обструкции и наличием (если имеются) признаков эмфиземы. Точно так же строится и терапия — на тех основаниях, которые есть для БА и для ХОБЛ.
Базисная фармакотерапия, включающая сочетание ИГКС с длительно действующими бронходилататорами (ДДБА, ДДмХБ или ДДБА + ДДмХБ). Если мы при БА можем в некоторых случаях проводить монотерапию ИГКС, то у пациентов с синдромом перекреста не должна проводиться монотерапия ИГКС или бронходилататорами. Всегда ИГКС должны сочетаться с длительно действующими бронходилататорами.
Другие важные стратегии лечения:
— отказ от курения,
— легочная реабилитация,
— вакцинация,
— лечение сопутствующих заболеваний.
Поскольку синдром перекреста отличается от «чистой», изолированной БА тем, что там присутствует ХОБЛ, программа реабилитации имеет чрезвычайно важное значение, нередко не менее значимое, чем фармакотерапия.Для успеха реабилитационных программ при ХОБЛ важное значение имеет адекватная медикаментозная терапия. В исследованиях было показано положительное влияние комбинации формотерола/будесонида на переносимость физической нагрузки и уровень одышки.
Еще один важный аспект – бронхопротективный эффект длительно действующих бронхолитиков. Обычно мы говорим о бронходилатирующих эффектах как об очень значимых. Но важна роль препарата и в профилактике возникновения бронхоконстрикции под воздействием различных факторов внешней среды. Длительно действующим бета-2-агонистам (ДДБА) и длительно действующим холинолитикам (ДДХЛ) отдается предпочтение в терапии таких пациентов.
Следует сказать несколько слов про использование холинергических препаратов при ХОБЛ, синдроме перекреста и при БА. Есть схема — участие парасимпатических нервных систем в БА и ХОБЛ, которую приводит в своих работах профессор Барнес (рис. 10). Здесь показана не только роль парасимпатической нервной системы в формировании повышенного тонуса парасимпатикуса и при бронхоспазме, но и то, что холинергические рецепторы, являются воспринимающим рецепторным полем для разных ирритарных воздействий, что ведет еще к более мощному формированию аксон-рефлекса с выбросом ацетилхолина и последующим каскадом реакций. С этим связаны предпосылки к использованию антихолинергических препаратов при бронхообструктивной патологии:
Есть схема — участие парасимпатических нервных систем в БА и ХОБЛ, которую приводит в своих работах профессор Барнес (рис. 10). Здесь показана не только роль парасимпатической нервной системы в формировании повышенного тонуса парасимпатикуса и при бронхоспазме, но и то, что холинергические рецепторы, являются воспринимающим рецепторным полем для разных ирритарных воздействий, что ведет еще к более мощному формированию аксон-рефлекса с выбросом ацетилхолина и последующим каскадом реакций. С этим связаны предпосылки к использованию антихолинергических препаратов при бронхообструктивной патологии:
• Холинергические механизмы участвуют в патогенезе ХОБЛ, БА, СПАХ
• Влияние на ФВД (бронходилатация, снижение гиперинфляции)
• Снижают бронхиальную гиперреактивность и оказывают бронхопротективный эффект (метахолиновая бронхоконстрикция, β-блокаторы)
• Регулярное использование уменьшает гипертрофию гладких мышц дыхательных путей (профилактика ремоделирования бронхов и вторичной бронхиальной гиперреактивности)
• Уменьшение холинергически-опосредованной секреции слизи и ингибирование гиперплазии бокаловидных клеток и слизистых желез
• Снижение нейтрофильного ответа в нижних дыхательных путях
• Снижение экспрессии провоспалительных генов в гладких мышцах дыхательных путей и бронхиального эпителия
• Оказывает стероид-спаринг эффект у больных БА
Напоминаю вам в связи с этим о препарате гликопирроний (Сибри), который выпускается и используется с помощью ингалятора Бризхалер. Бронходилатационный эффект у Сибри сохраняется через 24 часа. Прием гликопиррония на протяжении года улучшает ОФВ1, снижается частота обострений, увеличивается время переносимости физических нагрузок. Кроме того, Сибри, как и Форадил, обладает бронхопротективным эффектом.
Бронходилатационный эффект у Сибри сохраняется через 24 часа. Прием гликопиррония на протяжении года улучшает ОФВ1, снижается частота обострений, увеличивается время переносимости физических нагрузок. Кроме того, Сибри, как и Форадил, обладает бронхопротективным эффектом.
В заключение хочется сказать несколько слов о средствах доставки препаратов. Мало создать хорошую молекулу — нужно суметь эффективно доставить ее в дыхательные пути. Два ингалятора — порошковый и аэрозольный, бризхалер и аэролайзер — отличаются, во-первых, наличием низкого сопротивления, что важно для реализации хорошего вдоха. Ингаляторы с низким сопротивлением позволяют пациенту произвести эффективный маневр дыхания. Это приводит к тому, что депозиция легочной ткани возрастает существенно, а нелегочной — уменьшается. Значит, улучшается профиль безопасности.
Кроме того, важна возможность контроля пациентами своих действий. В данном случае при использовании этих ингаляторов пациенты:
• слышат, как вращается капсула при вдохе,
• чувствуют сладковатый привкус препарата (как при ингаляции формотерола, так и будесонида),
• видят насколько опорожнилась капсула после вдоха (при необходимости можно повторить вдох).
Таким образом, пациенты полностью контролируют процесс ингаляции.
Основным принципом лечения больных астмой при СПАХ остается ступенчатый подход: увеличивается объем терапии при снижении контроля или наличии факторов риска обострений, уменьшается — при достижении и сохранении приемлемого контроля заболевания в каждом конкретном случае.
Комбинация будесонид/формотерол в сочетании с ДДмХБ (и в частности, с гликопирронием) является эффективным выбором при ведении больных БА и ХОБЛ (СПАХ).
Гость симпозиума – сотрудник университетской клиники им. К.Г. Каруса (г. Дрезден, Германия), профессор Герт Хёффкен выступил с докладом «Опыт ведения пациентов с астмой в Германии»:
Как и во всем мире, врачи в Германии используют стандарты GINA. Однако у нас есть свои национальные рекомендации, которые немного отличаются от международных. Например, у нас используется всего 4 ступени терапии БА, а не пять. 4 и 5 ступени объединены в одно целое. И препараты тиотропия, согласно нашим рекомендациям, назначаются уже на 4 ступени терапии.
Рациональное мышление, принципами которого мы, конечно же, руководствуемся при лечении пациентов, это, во-первых, определение фенотипа БА. Эндотип определить сложнее — это более экспериментальный подход, но фенотипы (ТН-2 опосредованная астма или ТН-1) определяются обязательно. Используются биомаркеры — такие как уровень IgE, уровень эозинофилов, измеряется уровень оксида азота в выдыхаемом воздухе, проводятся тесты по переостину. Но, насколько мне известно, в России они не зарегистрированы.
Во-вторых, естественно проводится сбор анамнеза, выявление триггеров и патогенов, определение гиперчувствительности. Требуется объяснить пациенту, в чем конкретно заключается его заболевание, что его провоцирует. У нас есть масса образовательных материалов, которые мы используем в общении с нашими больными.
Терапия БА начинается с ингаляционных кортикостероидов. ИГКС – краеугольный камень в терапии БА, особенно в комбинации с ДДБА. Мы имеем широкий выбор ингаляционных кортикостероидов. Но какого-то определенного препарата, указываемого в национальных рекомендациях, нет. Мы используем наиболее часто комбинацию из будесонида и формотерола или флутиказона с сальметеролом. Лично я предпочитаю первый вариант. Хочу сказать, что я полностью согласен с российскими коллегами: наличие одного прибора для доставки двух препаратов — один ингалятор с двумя компонентами — предпочтительнее двух разных. Ведь мы не должны забывать об удобстве для пациентов. Поэтому мы придерживаемся подхода комбинирования в одном устройстве двух компонентов. На рисунке 11 вы можете посмотреть результаты исследования «Фиксированные комбинации ИГКС/ДДБА vs монокомпоненты в 2 ингаляторах: взгляд пациентов», проведенного в 2013 г.
Но какого-то определенного препарата, указываемого в национальных рекомендациях, нет. Мы используем наиболее часто комбинацию из будесонида и формотерола или флутиказона с сальметеролом. Лично я предпочитаю первый вариант. Хочу сказать, что я полностью согласен с российскими коллегами: наличие одного прибора для доставки двух препаратов — один ингалятор с двумя компонентами — предпочтительнее двух разных. Ведь мы не должны забывать об удобстве для пациентов. Поэтому мы придерживаемся подхода комбинирования в одном устройстве двух компонентов. На рисунке 11 вы можете посмотреть результаты исследования «Фиксированные комбинации ИГКС/ДДБА vs монокомпоненты в 2 ингаляторах: взгляд пациентов», проведенного в 2013 г.
Реальная жизнь, как в Германии, так и в России, и во всем мире, естественно, отличается от того, что написано в рекомендациях экспертного заключения. Число пациентов с полностью контролируемой астмой или частично контролируемой составляет около 50% от общего количества страдающих этим заболеванием (рис. 12). Исследование, проведенное в Германии несколько лет назад с участием 483 тыс. пациентов, показало, что обеспеченность необходимыми препаратами (даже для базисной терапии), к сожалению, все еще невысокая. Кто-то из пациентов получает только КДБА, кто-то ДДБА как монотерапию, но это, конечно, неправильно. Вот почему такие мероприятия действительно важны для нас, чтобы мы могли услышать друг друга и обменяться опытом.
12). Исследование, проведенное в Германии несколько лет назад с участием 483 тыс. пациентов, показало, что обеспеченность необходимыми препаратами (даже для базисной терапии), к сожалению, все еще невысокая. Кто-то из пациентов получает только КДБА, кто-то ДДБА как монотерапию, но это, конечно, неправильно. Вот почему такие мероприятия действительно важны для нас, чтобы мы могли услышать друг друга и обменяться опытом.
Запись симпозиума: Маргарита Полякова
особенности этиологии, патогенеза, диагностики, фармакотерапии – тема научной статьи по клинической медицине читайте бесплатно текст научно-исследовательской работы в электронной библиотеке КиберЛенинка
УДК 616.248: 616.233-002: 616.233-007.271: 615.234: 615.217.34: 577.175.53
сочетание бронхиальной астмы и хронической обструктивной болезни лёгких: особенности этиологии, патогенеза, диагностики, фармакотерапии
Олег Михайлович Урясьев, Светлана Васильевна Фалетрова*, Людмила Владимировна Коршунова
Рязанский государственный медицинский университет им. И.П. Павлова, г. Рязань, Россия
И.П. Павлова, г. Рязань, Россия
Поступила 12.02.2016; принята в печать 16.02.2016.
Реферат DOI: 10.17750/KMJ2016-394
Бронхиальная астма и хроническая обструктивная болезнь лёгких — самые распространённые обструктивные заболевания респираторной системы. Хронической обструктивной болезнью лёгких страдают 230 млн человек, бронхиальной астмой — 300 млн человек в мире. Ежегодно от хронической обструктивной болезни лёгких умирают 200-300 человек в Европе и 2,74 млн населения в мире, от бронхиальной астмы — 250 тыс. человек в год. Социальная и экономическая значимость этих заболеваний определяют необходимость глубокого изучения их сочетания у одного пациента. Каждому заболеванию присущи свои фенотипы, но у 10-20% больных присутствуют признаки как хронической обструктивной болезни лёгких, так и бронхиальной астмы. Несмотря на чёткие диагностические критерии, в ряде случаев трудно провести границу между данными заболеваниями. Морфологической основой этих заболеваний является хроническое воспаление в бронхиальном дереве, нарушающее целостность эпителия, обусловливающее запуск бронхоконстрикторных реакций и приводящее к необратимой бронхиальной обструкции, присущей как тяжёлой бронхиальной обструкции, так и хронической обструктивной болезни лёгких. Однако стратегии лечения бронхиальной астмы и хронической обструктивной болезни лёгких имеют существенные различия, поэтому важно иметь чёткие диагностические критерии для выделения различных фенотипов, в том числе сочетанного фенотипа бронхиальной астмы и хронической обструктивной болезни лёгких. Рациональная стартовая терапия синдрома перекрёста бронхиальной астмы и хронической обструктивной болезни лёгких включает препараты, воздействующие на патогенетические механизмы обоих заболеваний, и представляет собой сочетание ингаляционных глюкокортикоидов с комбинированной бронходилатационной терапией — ß2-агонистами длительного действия и пролонгированными антихолинергическими препаратами.
Морфологической основой этих заболеваний является хроническое воспаление в бронхиальном дереве, нарушающее целостность эпителия, обусловливающее запуск бронхоконстрикторных реакций и приводящее к необратимой бронхиальной обструкции, присущей как тяжёлой бронхиальной обструкции, так и хронической обструктивной болезни лёгких. Однако стратегии лечения бронхиальной астмы и хронической обструктивной болезни лёгких имеют существенные различия, поэтому важно иметь чёткие диагностические критерии для выделения различных фенотипов, в том числе сочетанного фенотипа бронхиальной астмы и хронической обструктивной болезни лёгких. Рациональная стартовая терапия синдрома перекрёста бронхиальной астмы и хронической обструктивной болезни лёгких включает препараты, воздействующие на патогенетические механизмы обоих заболеваний, и представляет собой сочетание ингаляционных глюкокортикоидов с комбинированной бронходилатационной терапией — ß2-агонистами длительного действия и пролонгированными антихолинергическими препаратами.
Ключевые слова: бронхиальная астма, хроническая обструктивная болезнь лёгких, фенотип, синдром перекрёста бронхиальной астмы и хронической обструктивной болезни лёгких, воспаление.
COMBINATION OF ASTHMA AND CHRONIC OBSTRUCTIVE PULMONARY DISEASE: FEATURES OF ETIOLOGY, PATHOGENESIS, DIAGNOSIS, PHARMACOTHERAPY
O.M. Uryasev, S.V. Faletrova, L.V. Korshunova
Ryazan State Medical University named after academician I.P. Pavlov, Ryazan, Russia
Bronchial asthma and chronic obstructive pulmonary disease are the most common obstructive diseases of the respiratory system. 230 million people suffer from chronic obstructive pulmonary disease, from bronchial asthma — 300 million people worldwide. Annually 200-300 people in Europe and 2. 74 million of world population die from chronic obstructive pulmonary disease, from asthma — 250 thousand people a year. The social and economic significance of these diseases determine the need for in-depth study of their combination in the same patient. Each disease has its own phenotypes, but in 10-20% of patients, there are symptoms of both chronic obstructive pulmonary disease and asthma. In spite of clear diagnostic criteria, in some cases it is difficult to distinguish these diseases. Morphological basis of these diseases is a chronic inflammation in the bronchial tree that causes damage to the epithelial continuity that initiates bronchoconstrictive reaction and leads to irreversible airway obstruction attributable for both severe bronchial obstruction and chronic obstructive pulmonary disease. However, the treatment strategy of bronchial asthma and chronic obstructive pulmonary disease has significant differences, it is important to have a clear diagnostic criteria to distinguish different phenotypes, including those of combined phenotype of asthma and chronic obstructive pulmonary disease.
74 million of world population die from chronic obstructive pulmonary disease, from asthma — 250 thousand people a year. The social and economic significance of these diseases determine the need for in-depth study of their combination in the same patient. Each disease has its own phenotypes, but in 10-20% of patients, there are symptoms of both chronic obstructive pulmonary disease and asthma. In spite of clear diagnostic criteria, in some cases it is difficult to distinguish these diseases. Morphological basis of these diseases is a chronic inflammation in the bronchial tree that causes damage to the epithelial continuity that initiates bronchoconstrictive reaction and leads to irreversible airway obstruction attributable for both severe bronchial obstruction and chronic obstructive pulmonary disease. However, the treatment strategy of bronchial asthma and chronic obstructive pulmonary disease has significant differences, it is important to have a clear diagnostic criteria to distinguish different phenotypes, including those of combined phenotype of asthma and chronic obstructive pulmonary disease. Rational starting therapy of asthma and chronic obstructive pulmonary disease overlap syndrome includes drugs acting on the pathogenic mechanisms of both diseases, and is a combination of inhaled corticosteroids with combined bronchodilator therapy — long-acting ß2-agonists and long-acting anticholinergics.
Rational starting therapy of asthma and chronic obstructive pulmonary disease overlap syndrome includes drugs acting on the pathogenic mechanisms of both diseases, and is a combination of inhaled corticosteroids with combined bronchodilator therapy — long-acting ß2-agonists and long-acting anticholinergics.
Keywords: bronchial asthma, chronic obstructive pulmonary disease, phenotype, asthma and chronic obstructive pulmonary disease overlap syndrome, inflammation.
Бронхиальная астма (БА) и хроническая обструктивная болезнь лёгких (ХОБЛ) — два самостоятельных, наиболее часто встречающихся заболевания респираторной системы. По данным Всемирной организации здравоохранения (ВОЗ), в мире ХОБЛ страдают 230 млн человек, из них 11,8% мужчин и 8,55% женщин старше 40 лет [33]. Ежегодно от ХОБЛ умирают 200-300 человек в Европе и 2,74 млн населения в мире [9]. По результатам «Исследования глобального ущерба
Адрес для переписки: faletrova@yandex. ru 394
ru 394
от заболеваний», данная нозология к 2020 г. выйдет на 3-е место, а к 2030 г. — на 4-е место среди причин смерти (в 1990 г. ХОБЛ занимала 6-е место в структуре смертности) [35].
В течение длительного периода ХОБЛ затрагивала преимущественно мужчин. В настоящее время во всём мире возросло потребление табака женщинами, что привело к росту заболеваемости ХОБЛ среди женщин.
В мире около 300 млн человек страдают БА — каждый 20-й житель планеты. Распространённость этого заболевания в разных странах мира
колеблется от 1 до 18% [10, 26]. По данным ВОЗ, летальность от БА составляет 250 тыс. человек в год. Исходя из результатов аналитико-прогнос-тических исследований, не исключено, что при стремительных темпах урбанизации к 2025 г. БА разовьётся дополнительно у 100-150 млн человек [16].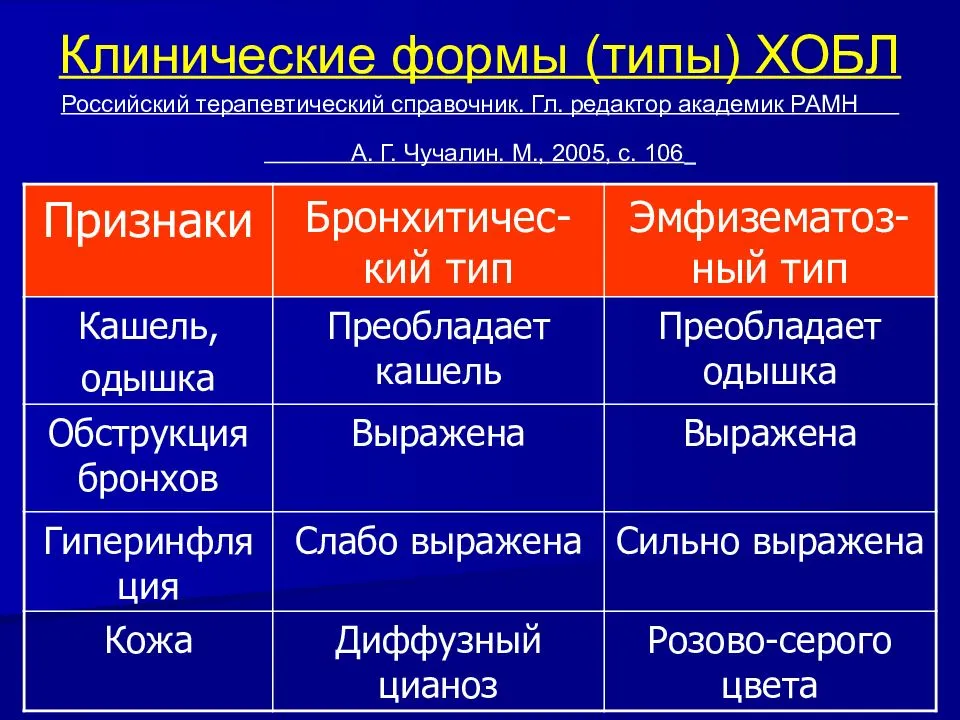
Социальная и экономическая значимость ХОБЛ и БА определяют необходимость глубокого изучения их сочетания у одного пациента. Несмотря на чёткие диагностические критерии, в ряде случаев трудно провести границу между этими заболеваниями [23, 32]. В 2014 г. впервые выделен синдром перекрёста БА и ХОБЛ (СПБАХ), что дало врачам возможность избегать как гипер- так и гиподиагностики БА и ХОБЛ.
У пациента может быть атопический фенотип, пароксизмальные симптомы, но при этом присутствовать прогрессирующая обструкция, не полностью обратимая на фоне адекватной терапии, низкая эффективность стероидных противовоспалительных препаратов, в анамнезе длительный стаж курения, воздействие других факторов риска. В этой ситуации тяжёлая форма БА может быть по ошибке расценена как ХОБЛ [3, 8, 11].
С другой стороны, у 20-30% больных ХОБЛ отмечают хороший ответ на терапию глюкокор-тикоидами (ГК), выявляют эозинофилию мокроты — признаки, типичные для БА [19, 27]. Возможна выраженная вариабельность скорости снижения объёма форсированного выдоха за первую секунду (ОФВ1). По результатам исследования «Evaluation of COPD Longitudinlly to Identifiy Predicyive Surrogate Endpoint (ECLIPSE)», у 50% пациентов с ХОБЛ ОФВ1 не отличается от возрастной нормы, при этом нередко ставят диагноз БА [22, 43, 45]. Именно в этих ситуациях правомочен диагноз сочетания БА и ХОБЛ.
Возможна выраженная вариабельность скорости снижения объёма форсированного выдоха за первую секунду (ОФВ1). По результатам исследования «Evaluation of COPD Longitudinlly to Identifiy Predicyive Surrogate Endpoint (ECLIPSE)», у 50% пациентов с ХОБЛ ОФВ1 не отличается от возрастной нормы, при этом нередко ставят диагноз БА [22, 43, 45]. Именно в этих ситуациях правомочен диагноз сочетания БА и ХОБЛ.
Пациенты с данным синдромом имеют «своё лицо», свой особый фенотип. Термин «фенотип» предложен одним из основателей современной генетики, датским биологом В. Иогансеном в 1909 г. Фенотип — видимые характеристики организма, обусловленные взаимодействием его генетической составляющей и факторов внешней среды. Фенотипизация в медицине — это оптимизация диагностики, лечения и профилактики [18]. Как ХОБЛ, так и БА присущи разные фенотипы, которые могут модифицироваться по мере увеличения длительности заболевания.
БА изучают много веков. Термин «бронхиальная астма» был введён Гиппократом (460-377 гг. до н.э.), он объединял все патологические состояния, сопровождающиеся удушьем. Греческий врач Артей Каппадокийский (I—II век до н.э) около 1800 лет назад классифицировал БА и предпринял первую попытку выделить фенотипы этой болезни.
В настоящее время в повторных редакциях согласительного документа по диагностике и лечению БА (GINA — от англ. Global Initiative for Asthma) обобщён накопленный материал, касающийся всех аспектов этой болезни [29]. Выделено
множество фенотипов заболевания.
У ряда больных БА характеризуется низким уровнем контроля течения заболевания, более высокой частотой обострений, сниженным ответом на ингаляционные ГК, большей потребностью в препаратах неотложной помощи. При этом возникает необратимая бронхиальная обструкция, связанная с длительно существующим воспалением и ассоциированная со структурными изменениями бронхолёгочной системы, на основании чего выделяют фенотип тяжёлой БА (нейтрофильная, курильщика, резистентная к ГК) [16, 17, 40, 42].
Фенотипирование ХОБЛ также имеет свою историю. Длительное время эту болезнь называли хроническим обструктивным бронхитом или эмфиземой лёгких — в зависимости от преобладания тех или иных симптомов. В последние десятилетия XX века учёные всего мира решили называть его везде одинаково — ХОБЛ.
В 2001 г разработана программа глобальной стратегии диагностики, лечения и профилактики этого заболевания (GOLD — от англ. Global Initiative for Chronic Obstructive Lung Disease). Данный документ пересматривали в 2006, 2011, 2013 и 2014 гг. [19]. Уже в первой редакции GOLD было выделено три основных фенотипа ХОБЛ: бронхитический, эмфизематозный, смешанный.
В последующем были описаны другие фенотипы этого заболевания, отличающиеся выраженностью и характером воспаления, наличием сопутствующих заболеваний, ответом на терапию, вариабельностью бронхиальной обструкции [30]. В 2014 г. выделен фенотип сочетания БА и ХОБЛ, определено наличие ряда общих черт при различных механизмах развития данных заболеваний [9].
Согласно определению, приведённому в GINA, БА представляет собой гетерогенное заболевание, которое, как правило, характеризуется наличием хронического воспаления дыхательных путей. Оно определяется по наличию в анамнезе таких симптомов, как свистящие хрипы, одышка, вариабельное ограничение скорости воздушного потока на выдохе [10].
В GOLD редакции 2014 г. ХОБЛ определяется как заболевание, которое можно предотвратить и лечить, характеризующееся персистирующим ограничением скорости воздушного потока, которое обычно прогрессирует и связано с повышенным хроническим воспалительным ответом лёгких на действие патогенных частиц и газов [9].
Морфологической основой обоих заболеваний является хроническое воспаление в бронхиальном дереве, нарушающее целостность эпителиального слоя, обусловливающее запуск бронхоконстрикторных реакций, приводящее к необратимой бронхиальной обструкции, присущей как тяжёлой БА, так и ХОБЛ [15, 29]. При этом характер воспаления при БА и ХОБЛ имеет принципиальные различия.
При БА триггером становится сенсибилизирующее вещество. Воспалительный процесс охватывает все дыхательные пути, но физиологи-
ческие последствия особенно ярко проявляются в бронхах среднего калибра, без распространения на интерстициальную ткань и паренхиму лёгких. Воспаление характеризуется аллергическим, Th3-обусловленным фенотипом, то есть инфильтрацией эозинофилами и Т-хелперами 2-го типа. У куряшдх пациентов и пожилых больных при тяжёлой БА возможна нейтрофильная инфильтрация стенок бронхов (не связанный с Th3 фенотип). Ограничение проходимости дыхательных путей может быть полностью или не полностью обратимо [10, 27].
При ХОБЛ триггером бывает раздражающее вещество. Воспаление локализовано в мелких дыхательных путях, распространяется на интер-стициальную ткань и паренхиму лёгких, приводит к деструкции эластического каркаса стенок альвеол и формированию эмфиземы лёгких. Происходит аккумуляция нейтрофилов, цитотокси-ческих СD8+ Т-клеток и макрофагов [2, 6, 27]. У некоторых пациентов с ХОБЛ возможен смешанный характер воспаления с повышением уровня эозинофилов [9]. Ограничение проходимости дыхательных путей частично обратимо или необратимо.
Можно выделить общие факторы риска развития БА и ХОБЛ [9, 10, 14, 31].
I. Внутренние:
1) генетически детерминированная бронхиальная гиперреактивность;
2) нарушение роста и развития плода во время беременности;
II. Внешние:
1) ингаляционные воздействия — табачный дым, профессиональные вредности, загрязнение воздуха внутри и снаружи помещений, атмосферные поллютанты;
2) инфекции.
Особое внимание следует уделить воздействию табачного дыма. Среди лиц с неконтролируемой БА высока доля активных курильщиков. У курящего астматика риск развития ХОБЛ выше, чем у некурящего, хотя суммарная интенсивность курения ниже по сравнению с людьми, страдающими только ХОБЛ [39].
В настоящее время накоплены данные, указывающие на то обстоятельство, что патофизиология тяжёлой БА связана с малыми дыхательными путями. У курящих больных БА и у пациентов с сочетанием БА и ХОБЛ присутствуют выраженное воспаление и дисфункция малых дыхательных путей. По мере прогрессирования заболевания происходит поражение мельчайших бронхов, возникают структурные изменения в паренхиме, приводящие к гиперинфляции [7, 17].
Кроме того, табачный дым — источник окси-дантов. Оксидативный стресс вызывает необратимые повреждения структур паренхимы лёгких вследствие дисбаланса в системе «протеолиз-ан-типротеолиз», приводит к гиперинфляции, нарушает местный иммунный ответ. Эти механизмы составляют основу развития ХОБЛ и способствуют формированию необратимого компонента об-396
струкции при БА [14, 28, 36].
Сочетанные формы БА и ХОБЛ чаще всего развиваются при суммировании нескольких факторов риска, наиболее значимые из которых — курение, неадекватная терапия БА в дебюте заболевания, рецидивирующие респираторные инфекции. Однако реализация каждого отдельно взятого фактора зависит от генотипа пациента [31].
Схожесть симптомов зачастую вызывает трудности в диагностике и дифференциальной диагностике, особенно у курящих и пожилых пациентов, и как следствие — неверное определение тактики лечения больных.
Установлено, что в среднем у 10-20% больных присутствуют признаки и ХОБЛ, и БА, что становится взаимоотягощающим фактором, приводящим к большей выраженности симптомов заболеваний. Сочетание БА и ХОБЛ проявляется более частыми и тяжёлыми обострениями, низким качеством жизни, прогрессирующим снижением ОФВ1, не полностью обратимой обструкцией, прогрессирующим снижением толерантности к физической нагрузке, преимущественно ней-трофильным воспалением в слизистой оболочке бронхов, снижением эффективности ГК, увеличением потребности в Р2-агонистах по сравнению с пациентами с ХОБЛ без БА такого же возраста с аналогичной степенью бронхиальной обструкции [16, 28, 33, 39].
В связи с различиями в диагностических критериях БА и ХОБЛ и отсутствием «золотого стандарта» в дифференциальной диагностике данные о распространённости сочетания этих двух заболеваний у одного пациента варьируют в широких пределах. Частота сочетания БА и ХОБЛ колеблется от 12,1 до 55,2% среди пациентов с ХОБЛ и от 13,3 до 61,0% среди пациентов с БА [44, 46]. Наличие признаков обеих нозологических форм вызывает трудности в диагностике и определении лечебной тактики.
Для более детального изучения особенностей течения сочетания ХОБЛ и БА разработаны рекомендации по ведению данной категории больных [39].
В 2014 г. был обозначен новый термин — ACOS (от англ. Asthma-CORD Overlap Syndrome — синдром перекрёста БА и ХОБЛ). Научными комитетами GINA и GOLD на основании обзора литературы и соглашения экспертов разработан документ «Диагностика заболеваний с хроническим ограничением воздушного потока: БА, ХОБЛ и СПБАХ», в котором дано определение СПБАХ, доступно сформулированы критерии диагностики БА, ХОБЛ и СПБАХ и отражены подходы к тактике ведения больных [4, 12].
СПБАХ характеризуется персистирующим ограничением воздушного потока, рядом признаков, ассоциированных с БА, и рядом признаков, ассоциированных с ХОБЛ. Следовательно, СПБАХ определяется чертами, характерными одновременно для БА и ХОБЛ [34].
Основные положения, касающиеся подхода к ведению больных СПБАХ можно представить следующим образом [4, 10, 34]:
— для СПБАХ характерны признаки БА и ХОБЛ;
— дифференциальная диагностика трудна у пациентов пожилого и старческого возраста, а также курильщиков;
— рекомендован поэтапный подход к постановке диагноза;
— начальную диагностику СПБАХ можно эффективно проводить на уровне медицинских учреждений первичного звена, по показаниям возможно направление пациента на специализированные исследования;
— стартовую терапию выбирают с учётом преобладающих симптомов в сторону того или иного заболевания.
Поэтапный подход к диагностике СПБАХ выглядит следующим образом [10, 34]:
1) выявление пациентов с хроническим заболеванием дыхательных путей;
2) дифференциальная диагностика БА, ХОБЛ и СПБАХ;
3) спирометрическое подтверждение хронического ограничения воздушного потока;
4) назначение стартовой терапии и оценка её эффективности;
5) направление на специализированные исследования при наличии показаний.
Главным критерием диагностики СПБАХ служит наличие симптомов, характерных для обоих заболеваний:
— диагноз БА в анамнезе, аллергия и семейный анамнез БА и/или воздействие патогенных частиц;
— начало заболевания в возрасте 40 лет и старше;
— персистенция и вариабельность симптомов со стороны органов дыхания;
— отсутствие полной обратимости ограничения воздушного потока, улучшение показателей внешнего дыхания после бронходилататоров до отношения ОФВ1 к форсированной жизненной ёмкости лёгких <0,7;
— неуклонное прогрессирование заболевания, уменьшение выраженности симптомов на фоне лечения;
— изменения на рентгенограмме, характерные для ХОБЛ.
Если три признака и более свидетельствует в пользу как БА, так и ХОБЛ, то высока вероятность СПБАХ [4, 10, 24, 34].
Чаще всего ХОБЛ развивается на фоне БА. Это, как правило, больные в возрасте старше 40 лет. В анамнезе у таких пациентов — длительное воздействие факторов риска: курение, профессиональные и бытовые вредности [19]. Заболевание при смешанном фенотипе протекает тяжелее, обострения развиваются чаще, чем при изолированных БА и ХОБЛ, снижается эффективность ГК, неуклонно прогрессирует дыхательная недостаточность [38].
Присоединение БА к ХОБЛ происходит реже. В данном случае появляются волнообразные симптомы, связанные с бронхиальной обструкцией, пароксизмальные ночные симптомы, повышается потребность в бронходилататорах [19].
Лежащий в основе БА и ХОБЛ воспалительный процесс определяет схожесть основных проявлений этих заболеваний в виде кашля, одышки, сухих хрипов. Однако подходить к выбору лекарственных препаратов следует с учётом морфологических особенностей воспаления при каждом заболевании [6].
При подозрении на наличие у больного СПБАХ лечение нужно проводить в соответствии с рекомендациями по лечению БА [10, 34]. Базисными препаратами для больных БА служат ингаляционные ГК в сочетании с Р2-агонистами длительного действия.
В основе развития СПБАХ лежит сочетание эозинофильного и нейтрофильного воспаления, что нередко становится причиной снижения эффективности ингаляционных ГК. Применение бронхолитических средств возможно только в комбинации с ингаляционными ГК, так как при использовании их в виде монотерапии они будут маскировать прогрессирующее воспаление и могут привести к неконтролируемому течению БА.
Препаратами первой линии при ХОБЛ служат бронходилататоры или их комбинация с ингаляционными ГК. Последние также не используют в виде монотерапии, так как воспаление при ХОБЛ рефрактерно к ГК [5, 9].
Важное место в терапии обоих заболеваний занимают ингаляционные бронходилата-торы длительного действия (Р2-агонисты) и длительно действующие антихолинергические препараты. Существует две основные фармакологические стратегии бронходилатации: прямая, осуществляемая посредством стимуляции Р2-адренорецепторов, и непрямая — вследствие ин-гибирования влияния ацетилхолина на мускари-новые рецепторы [20]. Необходимо разобраться в рациональности, обоснованности, принципах назначения бронхолитиков различных групп отдельно и в комбинации при СПБАХ.
Решая вопрос о выборе бронхолитическо-го средства, нужно учитывать распределение рецепторов к данному лекарственному препарату в бронхиальном дереве. Плотность Р2-адренорецепторов одинакова в крупных и малых дыхательных путях. м-Холинорецепторы имеют высокую плотность в крупных дыхательных путях и низкую в малых. Следует отметить, что вагусная иннервация малых дыхательных путей отсутствует, но м-холинорецепторы локализуются на всём протяжении трахеобронхиального дерева и активируются экстранейрональным аце-тилхолином. Этим обусловлен бронходилатаци-онный эффект антихолинергических препаратов на уровне проксимальных и дистальных воздухоносных путей [41].
Доказано, что при тяжёлой БА и ХОБЛ эффективность Р2-агонистов может снижаться
вследствие сокращения количества активных Р2-адренорецепторов и уменьшения их чувствительности (особенно у пожилых пациентов). Важной особенностью м-холинорецепторов является то обстоятельство, что чувствительность их не меняется с возрастом. При ХОБЛ обратимый компонент бронхиальной обструкции контролируется парасимпатическим отделом вегетативной нервной системы.
Применение комбинации Р2-агониста и м-холинолитика имеет преимущества с точки зрения фармакодинамики и фармакокинетики. Р2-Агонисты усиливают расслабление бронхов, вызванное м-холиноблокаторами. Последние в свою очередь уменьшают бронхоконстриктив-ное действие ацетилхолина, усиливая эффект Р2-агонистов [6]. Применение Р2-агониста совместно с холиноблокатором улучшает проходимость бронхов в большей степени, чем монотерапия.
До недавнего времени альтернативы Р2-агонистам длительного действия в комбинации с ингаляционными ГК не было. Фиксированная комбинация Р2-агонистов длительного действия и ингаляционных ГК рациональна при лечении БА. Однако следует помнить, что контроль заболевания, особенно при тяжёлой БА, достигается менее чем в половине случаев [12]. В одном европейском исследовании показано, что на фоне адекватной терапии частичный контроль заболевания был достигнут в 40,6%, неконтролируемое течение было отмечено в 17,8% случаев [37].
В ряде исследований последних лет установлена немаловажная роль холинергических механизмов в патогенезе БА. Доказано, что тонус парасимпатической нервной системы может повышаться вследствие воздействия табачного дыма, воспаления, инфекций [21]. Кроме того, доказано, что полиморфизм Р2-рецепторов может быть причиной неэффективности терапии адре-номиметиками [1, 13]. При БА с нейтрофильным фенотипом воспаления добавление к терапии хо-линолитика позволяет уменьшить выраженность бронхиальной обструкции и предотвратить развитие тяжёлых обострений.
Таким образом, рациональная стартовая терапия СПБАХ включает препараты, воздействующие на патогенетические механизмы БА и ХОБЛ, и представляет собой сочетание ингаляционных ГК с комбинированной бронходилатационной терапией — Р2-агонистами длительного действия и пролонгированными антихолинергическими препаратами [5, 9, 23, 34].
Определяя дозу препаратов, не следует забывать о принципах лекарственной терапии: чем больше признаков БА — тем больше должна быть доза ингаляционных ГК [4]. Исходя из этого, при наличии неопределённости относительно диагноза можно выбрать самый безопасный вариант лечения и продолжать диагностический поиск [34].
Кроме фармакологического лечения, необходимо использовать другие виды терапии, проводить профилактические мероприятия, которые 398
будут способствовать повышению контроля симптомов, предупреждению прогрессирования заболевания, улучшению качества жизни, снижению смертности: отказ от курения, лёгочная реабилитация, вакцинация, лечение сопутствующих заболеваний [10, 34].
Имея представления об особенностях течения сочетания БА и ХОБЛ, используя конкретные диагностических критерии, опираясь на рекомендации по ведению данной категории больных, можно достичь контроля над заболеванием, повысить качество жизни, снизить летальность.
ЛИТЕРАТУРА
1. Авдеев С.Н. Эффективность и безопасность препарата Спирива респимат при ХОБЛ и бронхиальной астме. Эффективная фармакотерап. Пульмонол. и ото-риноларингол. 2014; 1 (18): 16-28. [Avdeev S.N. Efficacy and safety of Spiriva Respimat in patients with COPD and bronchial asthma. Effektivnaya farmakoterapiya. Pul’monologiya i otorinolaringologiya. 2014; 1 (18): 1628. (In Russ.)]
2. Астафьева Н.Г., Гамова И.В., Удовиченко Е.Н. и др. Фенотип бронхиальной астмы с ожирением: клинические особенности, диагностика, лечение. Астма и аллергия. 2015; (1): 1-8. [Astaf’eva N.G., Gamova I.V., Udovichenko E.N. et al. Phenotype of asthma with obesity: clinical features, diagnosis, treatment. Astma i allergiya. 2015; (1): 1-8. (In Russ.)]
3. Белевский А.С. Основные сведения об этиологии, патогенезе и лечении бронхиальной астмы. Астма и аллергия. 2015; (1): 15-18. [Belevskiy A.S. Understanding the etiology, pathogenesis and treatment of bronchial asthma. Astma i allergiya. 2015; (1): 15-18. (In Russ.)]
4. Белевский А.С. Синдром перекрёста бронхиальной астмы и хронической обструктивной болезни лёгких (по материалам совместного документа рабочих групп экспертов GINA и GOLD). Практ. пульмонол. 2014; (2): 12-19. [Belevskiy A.S. Cross over syndrome of bronchial asthma and chronic obstructive pulmonary disease (based on a joint document of expert working groups and GINA GOLD). Prakticheskaya pul’monologiya. 2014; (2): 12-19. (In Russ.)]
5. Белевский А.С., Княжеская Н.П. Сочетание хронической обструктивной болезни лёгких и бронхиальной астмы — лечебные возможности. Астма и аллергия. 2014; (2): 13-16. [Belevskiy A.S., Knyazheskaya N.P. The combination of chronic obstructive pulmonary disease and asthma-treatment possibilities. Astma i allergiya. 2014; (2): 13-16. (In Russ.)]
6. Бродская О.Н. Беродуал — универсальный препарат для лечения обострений бронхиальной астмы и ХОБЛ. Астма и аллергия. 2014; (4): 21-23. [Brodskaya O.N. Berodual — universal drug for the treatment of exacerbations of asthma and COPD. Asthma i allergiya. 2014; (4): 21-23. (In Russ.)]
7. Будневский А.В., Бурлачук В.Т., Олышева И.А. и др. Возможности контроля над бронхиальной астмой: роль малых дыхательных путей. Пульмонология. 2011; (2): 101-109. [Budnevskiy A.V., Burlachuk V.T., Olysheva I.A. et al. Control over asthma: the role of small ап-ways. Pul’monologiya. 2011; (2): 101-109. (In Russ.)]
8. Визель А.А., Визель И.Ю. Бронхиальная астма и хроническая обструктивная болезнь лёгких. http://www. consillium-medicum.com (дата обращения: 15.01.2016). [Vizel’ A.A., Vizel’ I.Yu. Bronchial asthma and chronic obstructive pulmonary disease. http://www.consillium-
medicum.com (Access date: 15.01.2016). (InRuss.)]
9. Глобальная стратегия диагностики, лечения и профилактики хронической обструктивной болезни лёгких (GOLD), пересмотр 2014 г. Пер. с англ. под ред. А.С. Белевского. М.: Российское респираторное общество. 2014; 92 с. [Global strategy for the diagnosis, treatment and prevention of chronic obstructive lung disease (GOLD), revision of 2014. Russ Ed.: Global’naya strategiya diagnostiki, lecheniya i profilaktiki khronicheskoy obstruktivnoy bolezni legkikh (GOLD), peresmotr 2014 g. Transl. from English ed. by A.S. Belevskiy. Moscow: Russian respiratory society. 2014; 92 p. (In Russ.)]
10. Глобальная стратегия лечения и профилактики бронхиальной астмы (GINA), пересмотр 2015 г. Пер. с англ. под ред. А.С. Белевского. М.: Российское респираторное общество. 2015; 148 с. [Global strategy for the diagnosis, treatment and prevention of chronic obstructive lung disease (GOLD), revision of 2015. Russ Ed.: Global’naya strategiya lecheniya i profilaktiki bronkhial’noy astmy (GINA), peresmotr 2015 g. Transl. from English ed. by A.S. Belevskiy. Moscow: Russian respiratory society. 2015; 148 p. (In Russ.)]
11. Дробик О.С., Битеева Д.В. Неконтролируемая бронхиальная астма — варианты решения проблемы. Астма и аллергия. 2013; (4): 11-18. [Drobik O.S., Biteeva D.V. Uncontrolled asthma — solutions. Asthma i allergiya. 2013; (4): 11-18. (In Russ.)]
12. Зыков К.А., Агапова О.Ю., Бейлина В.Б. и др. Новые подходы к лечению пациентов с сочетанием бронхиальной астмы и ХОБЛ — фокус на пролонгированные м-холинолитики. РМЖ. 2014; (18): 18361842. [Zykov K.A., Agapova O.Ju., Beylina V.B. et al. New approaches to treating patients with a combination of bronchial asthma and COPD — focus on prolonged m-anticholinergics. Rossiyskiy meditsinskiy zhurnal. 2014; (18): 1836-1842. (In Russ.)]
13. Иванов А.Ф., Черняк Б.А. Эффективность тиотропия бромида в лечении пациентов с бронхиальной астмой. Пульмонология. 2014; (4): 112-116. [Ivanov A.F., Chernjak B.A. Effectiveness of tiotropium bromide in the treatment of patients with bronchial asthma. Pul’monologiya. 2014; (4): 112-116. (In Russ.)]
14. Княжеская Н.П. Терапия хронической обструктивной болезни лёгких: возможности индакатерола. Consil. med. 2014; 16 (11): 13-17. [Knyazheskaya N.P. COPD therapy: indacaterol possibilities. Consilium medicum. 2014; 16 (11): 13-17. (In Russ.)]
15. Лобанова Е.Г., Калинина Е.П., Кнышова В.В. и др. Особенности регуляции иммунного ответа у пациентов с коморбидным течением хронической обструктивной болезни лёгких и бронхиальной астмы. Пульмонология. 2014; (6): 5-10. [Lobanova E.G., Kalinina E.P., Knyshova V.V. et al. Features of the immune response regulation in patients with comorbidities of the chronic obstructive pulmonary disease and asthma. Pul’monologiya. 2014; (6): 5-10. (In Russ.)]
16. Ненашева Н.М. Бронхиальная астма. М.: Атмосфера. 2011; 95 с. [Nenasheva N.M. Bronkhial’naya astma. (Bronchial asthma.) M.: Atmosfera. 2011; 95 p. (In Russ.)]
17. Ненашева Н.М. Персонализированная терапия бронхиальной астмы: реалии и перспективы. Практ. пульмонол. 2013; (4): 12-18. [Nenasheva N.M. Personalized therapy for bronchial asthma: realities and perspectives. Prakticheskaya pul’monologiya. 2013; (4): 12-18. (In Russ.)]
18. Ненашева Н.М. Фенотипы бронхиальной астмы и выбор терапии. Практ. пульмонол. 2014; (2): 2-11. [Nenasheva N.M. Phenotypes of asthma and the choice of therapy. Prakticheskaya pul’monologiya. 2014; (2): 2-11.
(In Russ.)]
19. Овчаренко С.И., Лещенко И.В. Хроническая об-структивная болезнь лёгких. М.: ГОЭТАР-Медиа. 2016; 189 с. [Ovcharenko S.I., Leshchenko I.V. Khronicheskaya obstruktivnaya bolezn’ legkikh. (Chronic obstructive pulmonary disease.) Moscow: GEOTAR-Media. 2016; 189 p. (In Russ.)]
20. Синопальников А.И. Антихолинергические препараты в лечении хронической обструктивной болезни лёгких: фокус на тиотропий. Пульмонология. 2014; (6): 73-82. [Sinopal’nikov A.I. Anticholinergic drugs in the treatment of chronic obstructive pulmonary disease: focus on tiotropium. Pul’monologyja. 2014; (6): 73-82. (In Russ.)]
21. Терехова Е.П. Оптимизация бронхиальной астмы с использованием длительно действующего антихолинергического препарата тиотропия бромида. Consil. Med. Болезни органов дыхания. 2014; (Прил.): 16-22. [Terekhova E.P. Optimization of bronchial asthma using long-acting anticholinergic drug tiotropium bromide. Consilium Medicum. Bolezni organov dykhaniya. 2014; (Pril.): 16-22. (In Russ.)]
22. Трофименко И.Н., Черняк Б.А. Предикторы тяжёлой одышки у больных хронической обструктивной болезнью лёгких средней степени тяжести. Пульмонология. 2014; (2): 55-59. [Trofimenko I.N., Chernyak B.A. Predictors of severe breathlessness in patients with chronic obstructive pulmonary disease of moderate severity. Pul’monologiya. 2014; (2): 55-59. (In Russ.)]
23. Урясьев О.М. Бронхиальная астма и заболевания сердечно-сосудистой системы. Земский врач. 2015; (4): 5-13. [Uryasev O.M. Comorbidity of bronchial asthma and cardiovascular disorders. Zemskiy vrach. 2015; (4): 5-13. (In Russ.)]
24. Урясьев О.М., Рогачиков А.И. Роль оксида азота в регуляции дыхательной системы. Наука молодых. 2014; (2): 133-140. [Uryasev O.M., Rogachikov A.I. Role of nitric oxide in regulation of respiratory system. Nauka molodykh — Eruditio Juvenium. 2014; (2): 133-140. (In Russ.)]
25. Федосеев Г.Б., Трофимов В.И., Шапорова Н.Л. и др. В поисках истины: что такое бронхиальная астма? Пульмонология. 2015; 25 (1): 5-15. [Fedoseev G.B., Trofimov V.I., Shaporova N.L. et al. In search of the truth: what is asthma? Pul’monologiya. 2015; 25 (1): 5-15. (In Russ.)]
26. Ходюшина И.Н., Урясьев О.М. Изменения показателей гемодинамики у больных бронхиальной астмой. Рос. мед.-биол. вестн. им. акад. И.П. Павлова. 2011; (2): 146-152. [Khodyushina I.N., Uryasev O.M. Changes of hemodynamics in the patients bronchial asthma. Rossiyskiy mediko-biologicheskiy vestnik imeni akademika I.P. Pavlova. 2011; (2): 146-152. (In Russ.)]
27. Чучалин А.Г., Авдеев С.Н., Айсанов З.Р. и др. Федеральные клинические рекомендации по диагностике и лечению хронической обструктивной болезни лёгких. Пульмонология. 2014; (3): 15-35. [Chuchalin A.G., Avdeev S.N., Aysanov Z.R. et al. Federal clinical recommendations for diagnosis and treatment of chronic obstructive pulmonary disease. Pul’monologiya. 2014; (3): 15-35. (In Russ.)]
28. Шапорова Н.Л., Трофимов В.И., Пелевина И.Д. и др. Поздняя астма: особенности клиники и лечебной тактики в амбулаторных условиях. Врач. 2013; (2): 22-24. [Shaporova N.L., Trofimov V.I., Pelevina I.D. et al. Late-onset asthma: specific features of its clinical presentation and management tactics in outpatient settings. Vrach. 2013; (2): 22-24. (In Russ.)]
29. Шапорова Н.Л., Трофимов В.И., Марченко В.Н. Бронхиальная астма тяжёлого течения. Атмосфера.
Пульмонол. и аллергол. 2014; (4): 3-6. [Shaporova N.L., Trofimov V.I., Marchenko V.N. Severe asthma. Atmosfera. Pul’monologyja i allergologiya. 2014; (4): 3-6. (In Russ.)]
30. Шмелёв Е.И. Современные возможности коррекции одышки у больных хронической обструк-тивной болезнью лёгких. Пульмонология. 2013; (6): 79-84. [Shmelev E.I. Modern possibilities of correction of breathlessness in patients with chronic obstructive pulmonary disease. Pul’monologiya. 2013; (6): 79-84. (In Russ.)]
31. Шмелёв Е.И. Сочетание бронхиальной астмы и хронической обструктивной болезни лёгких. Международ. мед. ж. 2006; (3): 49-53. [Shmelev E.I. Combination of bronchial asthma and chronic obstructive pulmonary disease. Mezhdunarodnyy meditsinskiy zhurnal. 2006; (3): 49-53. (In Russ.)]
32. Bateman E.D., Reddel H.K., van Zyl-Smit R.N. et al. The asthma-COPD overlap syndrome: towards a revised taxonomy of chronic airways diseases? The Lancet Respir. Med. 2015; 3 (9): 719-728.
33. Buist A.S., McBurnie M.A., Vollmer W.M. et al. Internacional variationin in the prevalence of COPD (the BOLD Study): a population-based prevalence studi. Lancet. 2007; 370: 741-750.
34. Diagnosis of diseases of chronic airflow limitation: asthma, COPD and asthma-COPD overlap syndrome (ACOS). Global Strategy for Asthma Management and Prevention. 2014. http//www.jinasthma.org (access date: 15.01.2016).
35. Mathers C.D., Loncar D. Projections of global mortality and burden of disease from 2002 to 2030. PLoSMed. 2006; 3 (11): 442.
36. Matsumura Y. Inflammation induces glucocorticoid resistance in patients with bronchial asthma. Anti-Inflammatory Anthi-Allergy Agents Med. Chem. 2009; 8 (4): 377-386.
37. Minchtva R., Ekerljung L., Bjerg A. et al. Frequent
УДК 612.13: 616.124.3-007.61: 616.131-008.331.1-08-039.73
cough in unsatisfactory controlled asthma-results from the population-based. West Sweden Asthma Study Respiratory Research. 2014; 15: 79.
38. Miravitlles M., Soler-Cataluna J.J., Calle M. et al. Spanish COPD Guidelines (Ges EPOC): pharmacological treatment of stable COPD. Spanish Society of Pulmonology and Thoracic Surgery. Arch. Broncopneumol. 2012; 48 (7): 247-257.
39. Miravitlles M., Soriano J.B., Ancochea J. et al. Characterisation of the overlap COPD — asthma phenotype. Focus on physican activity and health status. Respir. Med. 2013; 107: 1053-1060.
40. Moore W.C., Meyers D.A., Wenzel S.E. et al. Identification of asthma phenotypes using claster analysis in the Severe Asthma Research Program. Am. J. Respir. Crit. Care. Med. 2010; 181: 315-323.
41. Nardini S., Camiciottoli G., Locicero S. et al. COPD: maximization of bronchdilation. Multidiscip. Respir. Med. 2014; (9): 50-60.
42. Skold C.M. Remodeling in asthma and COPD — differences and similarities Clin. Respir. J. 2010; 4 (Suppl. 1): 20.
43. Tashkin D.P. Variations in FEV1 decline over time in chronic obstructive pulmonary disease and implecayions. Curr. Opin. Pulm. Med. 2013; 19 (2): 116-124.
44. Tho N.V., Park H.Y., Nakano Y. Asthma-COPD overlap syndrome (ACOS): diagnostic challenge. Respirology. 2015; (Oct. 8): doi: 10.1111/resp.12653 [Epub ahead of print].
45. Vestbo J., Edwards L.D., Scanlon P.D. et al. Chandges in forced expiratory volume in I second over time in CPTD. N. Engl. J. Med. 2011; 365: 1184-1192.
46. Wurst K.E., Kelly-Reif K., Bushnell G.A. et al. Understanding asthma-chronic obstructive pulmonary disease overlap syndrome. Respir Med. 2016; 110 (Jan.): 1-11.
особенности нарушений внутрисердечной
гемодинамики у пациентов с хронической
обструктивной болезнью лёгких, осложнённой
развитием хронического лёгочного сердца, и способы их медикаментозной коррекции
Назипа Ермухамбетовна Айдаргалиева, Асель Жаугаштыевна Телеушева* Казахский национальный медицинский университет им. С.Д. Асфендиярова, г. Алматы, Казахстан
Поступила 06.10.2015; принята в печать 19.01.2016.
Реферат DOI: 10.17750/ЮШ2016-400
Хроническое лёгочное сердце — распространённая патология, в последние годы ставшая одной из ведущих причин инвалидности и смертности [15]. По литературным данным, две трети пациентов с хроническими обструк-тивными заболеваниями лёгких умирают в период от 15 мес до 5 лет после проявления декомпенсации кровообращения, которая занимает 3-е место после артериальной гипертензии и ишемической болезни сердца среди причин смерти в возрастной группе старше 50 лет [14]. Основным патогенетическим механизмом развития хронического лёгочного сердца считают лёгочную гипертензию [11]. Смертность у больных с хроническим лёгочным сердцем напрямую связана с уровнем систолического давления в лёгочной артерии. Так, согласно литературным данным, при систолическом давлении в лёгочной артерии от 30 до 50 мм рт.ст. 4-5-летняя выживаемость составляет 30%, а при уровне более 50 мм рт.ст. 5-летняя выживаемость равна нулю [1]. Таким образом, лёгочная гипертензия — фактор неблагоприятного прогноза. Кроме того, известно, что изменения, выявленные в правом желудочке на фоне лёгочной гипертензии, являются предикторами выживания у пациентов с хронической обструктивной болезнью лёгких [18]. Обзор литературных источников показал, что особенность гемодинамики у больных с хроническим лёгочным сердцем — увеличение давления в лёгочной артерии, приводящее к процессам ремоделирования обоих желудочков, в частности к расширению правых отделов сердца, нарастанию диастолической дисфункции как пра-
Адрес для переписки: astel79@mail.ru 400
Page not found | Східноєвропейський журнал внутрішньої та сімейної медицини
Журнал орієнтований на лікаря-практика і його потреби. Журнал грунтується на клінічній доказовій базі, алгоритмах діагностики, лікування та профілактики, призводить максимальну кількість корисного ілюстративного матеріалу, на сайті журналу наводяться відео лекції та тестові завдання, що дають можливість фахівцеві в режимі он-лайн перевірити і підвищити свої знання.
Особливість нашого журналу — практично-орієнтований підхід, а також впровадження дистанційних форм навчання у повсякденну діяльність лікаря.
Статті публікуються українською, російською та англій- ською мовами. Приймаються в першу чергу оглядові статті з актуальних проблем внутрішньої та сімейної медицини, які можуть бути використані для дистанційної освіти лікарів та підвищення їх післядипломної освіти.
Автор (або колектив авторів) усвідомлює, що несе первісну відповідальність за новизну і достовірність результатів наукового дослідження, що передбачає дотримання таких принципів: •автори статті повинні надавати достовірні результати проведених досліджень. Завідомо помилкові або сфальсифіковані затвердження неприйнятні.
На сайті журналу постійно оновлюються клінічні протоколи та рекомендації щодо внутрішній медицині. Оформіть передплату на оновлення на сайті заповнивши форму, наведену вище і Ви завжди будете отримувати електронну розсилку з оновленнями.
Ми завжди відкриті для діалогу і співпраці. Ми будемо дуже вдячні за поради та рекомендації, які Ви можете надсилати нам. Ми дуже сподіваємося, що це видання отримає підтримку як у лікарів, так і у працівників системи післядипломної освіти. Ми вдячні всім, хто співпрацює з нашим виданням, і всім читачам за інтерес до основних проблем сучасної медицини!
Засновником журналу є Харківська обласна організація «Асоціація лікарів загальної практики — сімейної медицини». Харківська медична академія післядипломної освіти: Кафедра терапії, ревматології та клінічної фармакології. Харківський національний медичний університет: кафедра внутрішньої медицини №3. Для кореспонденції: teraprevm@med.edu.ua
ФАРМАТЕКА » К вопросу о фенотипах хронической обструктивной болезни легких
Для хронической обструктивной болезни легких (ХОБЛ) характерна значительная гетерогенность клинической картины и прогрессирования заболевания. Существующие диагностические показатели (в частности, объем форсированного выдоха за первую секунду) не могут полностью описать эту гетерогенность, в связи с чем возникла необходимость ввести понятие фенотипа болезни. Выделение отдельных фенотипов ХОБЛ не только позволяет объяснить различное течение заболевания, но и может помочь в выборе наиболее адекватной для каждого пациента терапии, что в свою очередь позволит улучшить качество жизни и прогноз больных ХОБЛ.
В последние годы отмечается рост числа заболеваний органов дыхания, среди которых особое место занимает хроническая обструктивная болезнь легких (ХОБЛ), представляющая значительную медицинскую проблему [1]. ХОБЛ является одним из наиболее распространенных заболеваний и развивается у 4–6 % мужчин и 1–3 % женщин, а в возрасте старше 40 лет этот процент увеличивается и достигает 11,8 % у мужчин и 8,5 % у женщин [2]. Это заболевание отличается высокой смертностью и значительными затратами ресурсов здравоохранения [3]. По данным экспертов
ВОЗ (2004), смертность от ХОБЛ в мире в 2002 г. составила 2,7 млн случаев, а по перспективной оценке, к 2020 г. ХОБЛ займет третье место как причина смерти с показателем в 4,5 млн случаев в год [4].
В нашей стране также наблюдается значительный рост смертности от ХОБЛ, однако истинное ее значение, по всей видимости, у нас недооценивается, поскольку в России показатели смертности в 2–3 раза ниже таковых в США и Европе. То же относится к диагностике ХОБЛ в России: на сегодняшний день у нас зарегистрировано чуть больше 2,4 млн пациентов, страдающих ХОБЛ, но предполагается, что их реальное число может быть в 10–11 раз больше [5].
Давая определение ХОБЛ, следует подчеркнуть, что это хроническое воспалительное заболевание, экологически опосредованное, в развитии которого ведущая роль принадлежит курению и воздействию аэрополютантов (промышленных и бытовых). Заболевание характеризуется частично обратимым, а в дальнейшем необратимым нарушением бронхиальной проходимости, преимущественным поражением дистальных отделов дыхательных путей и паренхимы легких, формированием эмфиземы. Болезнь отличается неуклонно прогрессирующим характером и
выраженными системными эффектами, которые могут усугублять ее течение [4].
В последние годы системным эффектам ХОБЛ придается все большее значение. Механизмы, лежащие в основе системных проявлений, запускаются курением, гипоксемией, малоподвижным образом жизни и системным воспалением. Воздействие медиаторов воспаления, продуктов оксидативного стресса и, возможно, иных пока еще малоизученных факторов не ограничивается исключительно легочной тканью: они оказывают влияние и на другие органы, а также системы организма.
К основным системным проявлениям ХОБЛ относятся дисфункция скелетных мышц, остеопороз, снижение питательного статуса и, что особенно важно, сердечно-сосудистые нарушения. В первую очередь у пациентов с ХОБЛ поражается скелетная мускулатура, при этом больной теряет мышечную массу. Это приводит к еще большему ограничению при выполнении им физических нагрузок. Пациенты с ХОБЛ имеют сниженную плотность костной ткани и более высокий риск переломов, а присоединение сердечно-сосудистой патологии еще больше ухудшает качество их жизни.
Что касается диагностики этого заболевания, то основным диагностическим критерием ХОБЛ служит функциональный показатель, представляющий собой постбронходилатационное отношение объема форсированного выдоха за первую секунду к форсированной жизненной емкости легких – ОФВ1/ФЖЕЛ < 70 %, по которому (при наличии воздействия на пациента факторов агрессии) ХОБЛ и выделяют из большой группы бронхообструктивных заболеваний.
Показатель ОФВ1 является очень ценным диагностическим инструментом и основным параметром легочной функции у этой категории больных, в первую очередь потому, что установлена значимость ОФВ1 как объективного критерия состояния бронхиальной проходимости и, следовательно, степени выраженности бронхиальной обструкции [4]. По величине ОФВ1 оценивают степень тяжести (стадию) заболевания, по его значению судят о прогрессировании ХОБЛ, определяют риск смерти, дают оценку проводимой лекарственной терапии [6].
Вместе с тем степень обструкции, оцененная по ОФВ1, слабо коррелирует с выраженностью симптомов (т. е. с клиническими проявления заболевания), качеством жизни, связанным со здоровьем. Больные ХОБЛ также различаются по клиническим проявлениям и скорости прогрессирования заболевания. Наблюдение за пациентами показало, что у многих больных с умеренным течением ХОБЛ отмечается
быстрое снижение легочной функции, в то же время у пациентов с тяжелой и очень тяжелой стадиями ХОБЛ активность заболевания не нарастает [7]. Все это создает проблемы при попытке улучшить диагностику ХОБЛ и классификацию ее тяжести [8].
В целом для ХОБЛ характерна значительная гетерогенность клинической картины и прогрессирования заболевания. Показатель ОФВ1 не может полностью описать эту гетерогенность, но в настоящее время другой альтернативы не найдено, хотя идет ее поиск.
С учетом вышеизложенного возникла необходимость ввести понятие фенотипа болезни (MeiLan K. Han, 2010) [9] и попытаться установить его среди больных ХОБЛ при II–IV стадиях заболевания. Исходя из определения фенотипа как совокупности всех свойств и признаков особи на определенной стадии развития, сформировавшихся в результате взаимодействия генотипа с окружающей средой, MeiLan K. Han и соавт. (2010) дали следующее определение фенотипа ХОБЛ: “Единственный отличительный признак заболевания или их комбинация, которые описывают различия между пациентами ХОБЛ, как они соотносятся с клиническими параметрами (симптомами, обострениями, ответом на терапию, скоростью прогрессирования заболевания или смерти)”.
Поскольку взаимосвязь спирометрических показателей и клинических проявлений слаба, функциональные параметры не могут адекватно отражать социальное значение ХОБЛ или эффективность терапевтических вмешательств, большинство исследователей считают гораздо более важным придавать значение изменению показателей, ориентированных на пациента: симптомов, обострений, переносимости физических нагрузок, качества жизни [7].
Фенотипу отводится чрезвычайно существенная роль. С его помощью открывается возможность классифицировать пациентов ХОБЛ по различным терапевтическим и прогностическим подгруппам, улучшать наше понимание процесса течения болезни. Возможно, с помощью более глубокого фенотипирования ХОБЛ вообще может возникнуть представление о ХОБЛ как о заболевании, объединяющем множество синдромов. Уже в настоящее время хорошо известны два классических фенотипа больных ХОБЛ – эмфизематозный с преобладанием одышки и бронхитический с продуктивным кашлем и рецидивирующими респираторными инфекциями [10], которые можно выделить среди больных ХОБЛ при III и IV стадиях заболевания. Для эмфизематозного типа (таких пациентов называют “розовыми пыхтельщиками”) характерна выраженная легочная гиперинфляция, часто имеется снижение массы тела, преобладает дыхательная недостаточность, легочное сердце развивается в поздние сроки и смерть наступает позднее. При бронхитическом типе (“синие одутловатики”) легочная гиперинфляция выражена слабо, но резко выражена обструкция, легочное сердце развивается раньше, быстро декомпенсируется, и смерть может наступить в среднем возрасте.
Изучение особенностей течения ХОБЛ у женщин [11] позволило выделить следующие особенности течения болезни:
• заболевание имеет более короткий анамнез;
• развивается при меньшем стаже курения;
• протекает с более выраженной одышкой;
• отличается более низкой физической выносливостью;
• имеется меньше сопутствующих заболеваний;
• параметры качества жизни хуже, чем у мужчин;
• более высокая выживаемость.
Все вышеизложенное позволяет выделить и еще один фенотип течения ХОБЛ – “женский пол”. Обсуждается существование и других фенотипов, таких как ХОБЛ в сочетании с бронхиальной астмой, болезнь с быстрым прогрессированием, раннее начало ХОБЛ, заболевание с частыми обострениями и “молодые больные”, ХОБЛ и дефицит α1-антитрипсина и др.
В настоящее время проводятся исследования, включающие в т. ч. изучение фенотипов ХОБЛ. В первую очередь к ним относится ECLIPSE (Evaluation of COPD Longitudinally to Identify Predictive Surrogate End-point), одной из задач которого служит определение клинически значимых фенотипов среди популяций больных ХОБЛ с II–IV стадиями течения (по GOLD – Global Strategy of Chronic Obstructive Lung Disease) и параметров, которые прогнозируют прогрессирование заболевания в течение трех лет у клинически значимых групп пациентов с ХОБЛ [12]. Исследование ECLIPSE продолжается, но уже первые результаты доложены на XX Конгрессе ERS (European Respiratory Society, 2010) и опубликованы в печати [13].
Изучались следующие, потенциально значимые (на современном этапе) особенности фенотипов у больных ХОБЛ:
• частота и тяжесть обострения;
• скорость снижения ОФВ1;
• выраженность бронходилатационного ответа;
• наличие системного воспаления.
Изучение частоты и тяжести обострения в зависимости от стадии ХОБЛ показало стабильность этого показателя. Обострения становятся более частыми и более тяжелыми по мере увеличения тяжести ХОБЛ. Фенотип, имеющий частые обострения, ассоциируется с более плохим качеством жизни этих больных.
Предварительные выводы, касающиеся скорости снижения ОФВ1, показали, что этот показатель отличается выраженной вариабельностью в ответ на проводимое лечение. Отсюда следует, что, будучи маркером тяжести течения ХОБЛ (именно по значениям ОФВ1 происходит классификация
тяжести болезни), ОФВ1 не является маркером активности заболевания. По мнению J. Vestbo и соавт., высказанному в докладе на XX Конгрессе ERS (2010), понятие тяжести ХОБЛ отличается от понятия степени активности процесса. Скорость снижения ОФВ1 не является стойким фенотипическим признаком ХОБЛ, т. к. наряду с падением легочной функции у ряда пациентов не отмечается этого падения, а у некоторых имеет место даже ее улучшение. Изучая выраженность бронходилатационного ответа на большой выборке (1831 больной ХОБЛ), исследователи пришли к заключению, что этот показатель не является стойким маркером ХОБЛ и не может быть признан фенотипической характеристикой заболевания.
Большое место в исследовании ECLIPSE отводится изучению воспаления, потенциальной роли биомаркеров при ХОБЛ. Первые выводы, касающиеся системного воспаления, показали, что его явное наличие определяется не у всех пациентов. Выводы предварительны, т. к. исследование продолжается и роль системного воспаления по-прежнему активно изучается, но первые результаты уже могут внести некоторую смуту в представление о системных проявлениях ХОБЛ.
Выделение отдельных фенотипов ХОБЛ не только позволяет объяснить различные течения заболевания, но и может помочь в выборе наиболее адекватной для каждого пациента терапии, что в свою очередь поможет улучшить качество жизни и прогноз больных ХОБЛ.
1. Хроническая обструктивная болезнь легких (Федеральная программа). Практическое руководство для врачей. Издание 2-е переработанное и дополненное / Под ред. академика РАМН А.Г. Чучалина. М., 2004.
2. Buist AS, McBurnie, et al. International variation in the prevalence of COPD (the BOLD Study):a population-based prevalence study. Lancet 2007;370:741–50.
3. Celli BR, MacNee W, Agusti AG, et al. Standards for the diagnosis and treatment of patients with COPD: a summary of the ATS/ERS position paper.Eur Respir J 2004;23:932–46.
4. Global strategy for the diagnosis, management and prevention of chronic obstructive pulmonary disease: GOLD executive summary. Am J Respir Crit Care Med 2007;176(6):532–55.
5. Социально-экономическое бремя бронхиальной астмы и хронической обструктивной болезни легких в Российской Федерации. РРОФ “Качество жизни” НИИ КЭФФ, М., 2009.
6. Committee for proprietary medicinal products points to consider on clinical investigation of medicine products in the chronic treatment of patients with COPD. London, CPMP / EWP/1999;562–98.
7. Jones PW, Agusti A. Outcomes and markers of chronic obstructive pulmonary disease. Eur Resp J 2006;27:822–32.
8. Cazzola M, MacNee W, et al. Outcomes for COPD pharmacological trials: from lung function to biomarkers. Eur Resp J 2008;31:416–69.
9. Han MeiLan K, Agusti A, Calverely P, et al. Chronic obstructive pulmonary disease phenotypes: the future of COPD. Am J Respir Crit Care Med 2010;182(5):598–604.
10. Хроническая обструктивная болезнь легких /Под ред. А.Г. Чучалина. M., 2008. C. 568.
11. Овчаренко С.И., Капустина В.А. Хроническая обструктивная болезнь легких: особенности у женщин // Пульмонология 2009. № 2. C. 102–12.
12. Vestbo J, Anderson W, Coxson HO, et al. Evaluation of COPD Longitudinally to Identify Predictive Surrogate End-point (ECLIPSE). Eur Respir J 2008;31:869–73.
13. Agusti A, Calverely P, Celli B, et al. Characterization of COPD heterogeneity in the ECLIPSE cohort.
Respir Research 2010;11:122.
Дифференциальный взгляд на отношения генотип-фенотип
Front Genet. 2015; 6: 179.
Virginie Orgogozo
1 CNRS, UMR 7592, Institut Jacques Monod, Université Paris Diderot, Paris, France
Baptiste Morizot
2 Aix Marseille Université, CNRS 7304, CNRS Прованс, Франция
Арно Мартен
3 Кафедра молекулярной клеточной биологии, Калифорнийский университет, Беркли, Калифорния, США
1 CNRS, UMR 7592, Институт Жака Моно, Парижский университет Дидро, Париж, Франция
2 Экс Марсельский университет, CNRS, CEPERC UMR 7304, Экс-ан-Прованс, Франция
3 Кафедра молекулярной клеточной биологии, Калифорнийский университет, Беркли, Калифорния, США
Отредактировал: Сильвен Марчеллини, Университет Консепсьон , Чили
Рецензент: Jacob A.Теннессен, Государственный университет Орегона, США; Маргарида Матос, Лиссабонский университет, Португалия
* Для корреспонденции: Virginie Orgogozo, CNRS, UMR 7592, Institut Jacques Monod, Université Paris Diderot, 15 Rue Hélène Brion, 75013 Paris, France gro.puselamron@ozogogro.einigrivThis article был отправлен в раздел «Эволюционная и популяционная генетика» журнала «Границы в генетике»
Поступило в редакцию 24 марта 2015 г .; Принято 28 апреля 2015 г.
Авторские права © 2015 Orgogozo, Morizot and Martin.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (CC BY). Использование, распространение или воспроизведение на других форумах разрешено при условии указания автора (авторов) или лицензиара и ссылки на оригинальную публикацию в этом журнале в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение без соблюдения этих условий.
Эта статья цитируется в других статьях в PMC.Abstract
Интегративный взгляд на разнообразие и своеобразие в живом мире требует лучшего понимания сложной связи между генотипами и фенотипами. Здесь мы вновь подчеркиваем старую точку зрения, согласно которой отношения генотип-фенотип (GP) лучше всего рассматривать как связь между двумя различиями, одно на генетическом уровне, а другое на фенотипическом. На сегодняшний день преобладающее мнение в биологических исследованиях состоит в том, что несколько генов взаимодействуют с множеством переменных окружающей среды (такими как абиотические факторы, культура или симбионты), чтобы произвести фенотип.Часто проблема связывания генотипов и фенотипов формулируется в терминах карт генотипов и фенотипов, и такие графические представления неявно уводят нас от дифференциального взгляда на отношения GP. Здесь мы показываем, что дифференциальный взгляд на отношения GP является полезной объяснительной структурой в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды. В таких случаях уместно рассматривать отношения GP как различия, встроенные в различия. Размышление о различиях проясняет сравнение между экологическим и генетическим влиянием на фенотипы и помогает лучше понять связь между генотипами и фенотипами.
Ключевые слова: генотип , фенотип, генетика, комплексный признак, GxE, GxG
Введение
Иногда кажется, что мы забываем, что исходный вопрос генетики заключался не в том, что делает белок, а скорее в том, «что делает собаку такой. собака, мужчина человек »
(Noble, 2006)
Один из фундаментальных вопросов биологии — понять, что отличает людей, популяции и виды друг от друга. Концепция фенотипа , который соответствует наблюдаемым признакам индивидуума, была придумана в противовес генотипу , унаследованному материалу, передаваемому гаметами.С момента первоначального предположения, что генотипы и фенотипы образуют два принципиально разных уровня биологической абстракции (Johannsen, 1911), проблема заключалась в том, чтобы понять, как они соединяются друг с другом, как генотипы отображаются на фенотипах. За последние 15 лет более 1000 примеров изменений последовательности ДНК были связаны с естественными не вредными фенотипическими различиями между людьми или видами у эукариот (Martin and Orgogozo, 2013b). У человека каталог OMIM ® (онлайн-менделевское наследование в человеке, http: // omim.org /), объединяющая генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей, и в общей сложности 2493 опубликованных исследования геномных ассоциаций (GWAS) выявили множество участков в геноме, которые статистически связаны со сложными признаками (Welter и др., 2014). Поскольку обнаружение причинных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в скоплении данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые хотят попытаться оспорить свое нынешнее понимание фенотипов.Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается проницательной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
Гены как факторы различия
Мутации, выделенные из лабораторных штаммов, сыграли важную роль в понимании карты GP. По классической схеме мутация сравнивается с эталоном дикого типа, и ее фенотипические эффекты используются для определения функции гена. Эта структура часто приводит к семантическому сокращению: из генетического изменения, вызывающего вариацию в фенотипе, часто удобно ассимилировать соответствующий ген в качестве причинной детерминанты признака (Keller, 2010; , рисунок ).Часто можно встретить заголовки, выражающие эти упрощения, трубящие перед широкой аудиторией об открытии гена «долголетия» или «благополучия», которые приносят научную точность в жертву психологическому воздействию. Исходя из этого, следует ли ген, чья мутация является летальной, называть «геном жизни»? Эти чрезмерно упрощенные формулировки на самом деле означают, что вариация в данном гене вызывает вариацию в данном фенотипе (Dawkins, 1982; Schwartz, 2000; Waters, 2007). Фактически, сам по себе ген не может ни вызвать наблюдаемого фенотипического признака, ни быть необходимым и достаточным для появления наблюдаемых признаков.Гены нуждаются в клеточной среде, комбинированном действии множества других генов, а также в определенных физико-химических условиях, чтобы иметь наблюдаемый эффект на организмы ( Рисунок ). Например, каштановая пигментация волос у одного человека является не только продуктом генов, кодирующих ферменты, синтезирующие пигмент, но также и присутствием клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного солнечный свет (Liu et al., 2013).Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, тщетен, чтобы полностью ответить на широкий вопрос о том, что делает волосы коричневыми, что вызывает конкретную биологическую структуру или процесс в целом. Тем не менее, генетический редукционизм может быть совершенно подходящим для идентификации генетических локусов, изменение которых вызывает фенотипические различия ( Рисунок ). Разница и цвета волос между двумя людьми в некоторых случаях может быть связана с их генетическими различиями.Отметим, однако, что не все фенотипические изменения можно объяснить генетическими изменениями. Разница в цвете волос также может быть вызвана негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окрашивание волос, или сочетанием генетических и негенетических различий.
Схематическое изображение линий GP. (A) Традиционное представление в классическом генетическом редукционизме. (B) Интегративный взгляд на биологию развития. (C) Схема экспериментального подхода в генетике. (D) Схема экспериментального подхода в эволюционной генетике. (E) Один пример таксономически устойчивой взаимосвязи GP: SLC45A2 и внутривидовые различия в пигментации у тигров и кур. Взаимосвязи GP обозначены пунктирными линиями на панелях (D – E) .
Пока современная генетика находилась в зачаточном состоянии, Альфред Стертевант сформулировал вопрос о карте GP простыми словами: «Одна из центральных проблем биологии — это проблема дифференциации — как яйцо превращается в сложный многоклеточный организм? Это, конечно, традиционная главная проблема эмбриологии; но это также проявляется в генетике в форме вопроса: «Как гены производят свои эффекты?» (Стертевант, 1932).Долгое время некоторые генетики, возможно, думали, что они вскрывают морфогенетические механизмы, лежащие в основе формирования фенотипических признаков, в то время как их экспериментальный подход фактически обнаруживал гены, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. Д.) Приводят к фенотипические различия (сравните фиг. с фиг. ). Фактически, предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и бледного пигмента феомеланина), либо как неявную ссылку на другие возможности. (он коричневый, а не другого цвета).Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наш разум и наш язык часто путают объекты, вариации которых рассматриваются, с самими вариациями (Keller, 2010), и важно напомнить, что в генетике интересующие объекты (например, данный генотип, аллель или фенотип) заслуживают определения относительно к другому эталонному состоянию.
Таким образом, классический генетический редукционистский подход по своей сути неспособен выяснить все факторы, ответственные за наблюдаемые характеристики в живом мире (Stotz, 2012), но является мощным и актуальным методом анализа генетических рычагов наследственной фенотипической изменчивости.Сосредоточение внимания на фенотипических вариациях между людьми, а не на абсолютных признаках, присутствующих в отдельных организмах, является ключом к лучшему пониманию генетических причин фенотипического разнообразия.
GP Взаимосвязь между двумя уровнями вариации
Мышление в терминах различий делает очевидной абстрактную сущность, которая включает в себя как генетический, так и фенотипический уровни. Эта сущность состоит из вариации в генетическом локусе (два аллеля), связанного с ней фенотипического изменения (два различных фенотипических состояния) и их взаимоотношений (, рисунок ).Мы втроем называем совокупность этих элементов «гефе», но здесь мы просто называем это «отношениями генотип-фенотип» (отношения GP). Мы покажем, что отношения GP — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия.
Генетическая часть отношения GP
В современных базах данных аннотаций генома ген обычно определяется как участок нуклеиновых кислот, который транскрибируется и кодирует РНК или полипептид с известной или предполагаемой функцией (Gerstein et al. al., 2007). Генетический локус, лежащий в основе фенотипических различий, не обязательно является геном в строгом смысле слова; он может охватывать конкретную пару оснований, кодирующую область, регуляторную область cis- или распространяться на весь ген с его регулирующими областями cis или даже на кластер генов ( Таблица ). Как ранее отмечалось другими (Falk, 1984; Gilbert, 2000; Stern, 2000; Moss, 2003; Griffiths and Stotz, 2013), концепция гена в биологии развития и в современных базах данных аннотаций генома отличается от концепции гена в эволюционная биология.Здесь упор делается не на сам ген, как это определено в геномных базах данных, а на индивидуальное функциональное разделение генома на локусы, создающие различия. Генотипическая часть родства GP может принимать форму различных аллелей: отдельные кодоны, кодирующие разные аминокислоты, вставки / делеции в кодирующей последовательности белка, расходящиеся версии определенного регуляторного элемента cis-, наличие / отсутствие вставок транспозонов. , количество копий гена в генном кластере, подверженном структурным изменениям и т. д.В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, включая так называемую мусорную ДНК (Graur et al., 2015), присутствие которой не влияет на наблюдаемые характеристики организма, кроме репликации и, возможно, транскрибирования. Есть также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и которые больше не связаны с фенотипической изменчивостью.Например, генетические вариации в кодирующих областях связывания гистоновой ДНК могли быть важны на ранней стадии эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически значимых вариаций, кроме летальных мутаций. Таким образом, в геноме есть нуклеотидные участки, которые абсолютно необходимы для жизни, но сами не содержат жизнеспособных фенотипически значимых вариаций.
Таблица 1
Несколько примеров отношений GP.
сайтов кодирования гены Время цветения Тале кресс, масличный рапс и9 кодирующие области
Агути| Генетический локус | Фенотипический признак | Организмы | ||||
|---|---|---|---|---|---|---|
| Два кодирующих сайта в гене ABO HBGG | Группа крови A / B | |||||
| человек, шимпанзе -регуляторный элемент в гене лактазы | Способность переваривать молоко | Различные популяции людей | ||||
| Число дупликаций генов амилазы AMY1 или AMY2B | Способность переваривать крахмал Человек | Собака | ||||
| Наличие / отсутствие комплекса смежных генов, кодирующих каротиноидные десатуразы и каротиноидциклазы / синтазы | Способность продуцировать каротиноиды | Гороховая тля, паутинный клещ, галлицовая муха, нематода | ||||
| Цветовое зрение | Человек, китообразные, рыбы, буйвол ies | |||||
| Кодирующая область F 3’5’H | Цветочная пигментация | Соя, горох, однолетний флокс, картофель, Иохрома | ||||
| Кодирующая область FRIGIDA | ||||||
| Кодовая область BADh3 | Аромат | Соя, рис | ||||
| Кодовая область HMA3 | Допуск тяжелых металлов | Cress и | Thale cress и цис -регулирующие области миостатина | Размер мышц | крупный рогатый скот, овца, собака, свинья, лошадь, человек | |
| цис -регулирующие области в комплексе ахаэто-щиток | Количество и положение сенсорных щетинок | Fruitflies | ||||
| Cis -регулирующий элемент в pitx1 900 36 ген | Морфология таза | Рыба-колюшка | ||||
| Кодирующая область SLC45A2 | Пигментация глаз, волос и кожи | Человек, тигр, курица | ||||
| Кодирующая область McK волос и кожи, но не глаза | Человек, мышь, крупный рогатый скот, курица, морская свинка, лошадь, лиса, свинья, овца, собака, кролик, медведь, ягуар, ягуарунди, белка, птицы, песчаная ящерица | |||||
| Cis — регулирующие области tan и ebony | Образец пигментации | плодовые мухи | ||||
| Cis — регулирующие элементы в гене shavenbaby | Положение и количество трихомов | |||||
| Плод 15159 | Cis — регулирующие элементы в гене optix | Красный узор на крыльях бабочки | Длиннокрылый бугорок rflies | |||
| Кодирующая область гемоглобина альфа и бета гены цепи | Устойчивость к гипоксии | Человек, лама, крокодил, олень, мышь, водоплавающая птица | ||||
Кодирующая область35 Aceинсектициды | Картофельный жук, тля, комары, комнатная муха, плодовая муха, восточная плодовая муха | |||||
| Наличие / отсутствие вставки CypA в гене TRIM5a | Устойчивость к ретровирусу | Сова, старая обезьяна World monkeys | ||||
| Сайты кодирования в Nav1.4 Ген | Устойчивость к тетродотоксину или саксотоксину | Змеи, иглобрюх, моллюски | ||||
| Кодирующая область TAS2R38 | Чувствительность к горечи | Человек, шимпанзе, кодирующие регионы | ||||
| Пигментация кожи и шерсти | Человек, олень, мышь, крупный рогатый скот, свинья, собака, кошка, лошадь, лиса, домашняя лиса, перепел, овца | |||||
| Количество дупликаций гена переносчика глюкозы HXT6 | Выживание в среде с низким содержанием глюкозы | Дрожжи | ||||
| Количество дупликаций гена CCL3L1 | Восприимчивость к ВИЧ-инфекции и скорость прогрессирования СПИДа после инфицирования | Человек | ||||
| Регуляторный элемент Cis — WntA ген | Пигментация крыльев | Бабочки 90 166 |
Фенотипическая часть отношения GP
Фенотипический аналог отношения GP относится к разновидности вариации (цвет волос, уровень устойчивости к токсинам и т. Д.)), а не до состояния (светлые волосы, дегустатор фенилтиокарбамида и т.д .; Таблица ).
Фенотип, связанный с генетическим изменением, не обязательно ограничивается организмом, который является носителем генетической мутации. Например, различие между левыми и правыми спиралями раковин улитки Lymnaea peregra определяется одним генетическим локусом с материнским эффектом: генотип матери, но не самой особи, отвечает за направление наматывание оболочки (Boycott et al., 1931). В других случаях причинно-следственные генетические изменения связаны с бактериями-симбионтами: термостойкость тли может варьироваться от человека к человеку из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты также могут проявляться на более высоком уровне, чем организм, у которого есть генетические изменения (Dawkins, 1982), одним из примеров является социальная организация колонии муравьев (Wang et al., 2013).
Дифференциальная часть отношения GP
Как определено выше, отношение GP включает в себя генетическое различие и фенотипическое различие.Связь между различиями как на генетическом, так и на фенотипическом уровне довольно абстрактна и может соответствовать трем отчетливым различиям в живом мире: (# 1) различие между двумя репродуктивно изолированными таксонами (живыми или вымершими), (# 2) различие, разделяющееся внутри популяции, и (# 3) различие, впервые появившееся в процессе эволюции, между организмом, несущим предковый аллель / признак, и его прямым потомком, который развил новый аллель / признак. Следует отметить, что изменение фенотипа не всегда следует за появлением новой вызывающей мутации сразу же, но может появиться позже из-за единственного набора аллелей, сегрегированных в популяции.Например, новый фенотип редуцированных пластин брони появился в популяции пресноводной колюшки, когда рецессивный аллель EDA , уже присутствующий на скрытых уровнях, оказался в гомозиготном состоянии у одного человека (Colosimo et al., 2005; Jones et al., 2012). Главный концептуальный прогресс, сделанный Чарльзом Дарвином, состоял в том, чтобы связать вариации между особями внутри скрещивающейся группы (различие № 2) с вариациями между таксономическими группами в пространстве и времени (различие № 1; Lewontin, 1974a).
Отметим также, что определенные фенотипические изменения могут проявляться на уровне всего организма, когда «причинная» мутация сопровождается дополнительными соматическими мутациями, которые весьма вероятны. Например, у женщин, несущих аллель дикого типа и мутантный аллель BRCA1 , клетки могут продуцировать белки BRCA1 дикого типа, поскольку они несут одну копию аллеля BRCA1 дикого типа. Тем не менее, эти женщины имеют до 80% риска развития рака груди или яичников к 70 годам по сравнению с женщинами, несущими две копии дикого типа BRCA1 , из-за появления дополнительных вредных мутаций в диком типе BRCA1 . аллель в их соматических клетках груди (Narod and Foulkes, 2004).
Важно отметить, что различие GP всегда определяется относительно представляющей интерес популяции или таксона (Sober, 1988). В менее развитых с медицинской точки зрения странах люди, несущие две дефектные копии гена фенилаланингидроксилазы, имеют серьезные проблемы со здоровьем, включая судороги и умственную отсталость. Напротив, в большинстве развитых стран у таких людей диагностируется при рождении, и они имеют нормальную продолжительность жизни с нормальным умственным развитием благодаря диете с ограничением фенилаланина (Armstrong and Tyler, 1955).Таким образом, связь между GP, включающая дефектную мутацию фенилаланингидроксилазы, зависит от контекста: мутация связана с проблемами со здоровьем в менее развитых с медицинской точки зрения странах, но не в других странах. Этот пример показывает, что причинная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика для случая фенилаланингидроксилазы) в рамках предположения ceteris paribus о «прочих равных условиях».
Таким образом, отношения GP лучше всего рассматривать как отношения между двумя вариациями, одна на генотипическом уровне, а другая на фенотипическом уровне. Человеческий разум может разработать концепции возрастающей абстракции: концепции вещей (например, клетки), концепции изменения (например, эволюции) и концепции отношений (например, гомология; Cassirer, 1910; Simondon, 1968). Здесь концепция отношения GP устанавливает связь между двумя изменениями (генетическими и фенотипическими). В следующих абзацах мы покажем, что, по сравнению с использованием интуитивных представлений о вещах, этот обходной путь через усиление абстракции может оказаться более эффективным для лучшего понимания фенотипического разнообразия.
Некоторые современные представления о связи между генотипом и фенотипом неявно отвергают дифференциальную точку зрения
Выше мы утверждали, что дифференциальную точку зрения следует всегда иметь в виду, думая о связи между генотипами и фенотипами. GWAS, которые представляют собой наиболее популярный метод обнаружения геномных локусов, связанных со сложными признаками в популяциях, основаны на анализе различий (Visscher et al., 2012). Тем не менее, в текущих исследованиях дифференциальная точка зрения иногда неявно отвергается.Когда наблюдаются множественные факторы, влияющие на фенотипические признаки ( Рисунок ), дифференциальная точка зрения считается слишком упрощенной, и исследователи часто предпочитают снова сосредоточиться на фенотипах отдельных людей, не связывая их явным образом с фенотипическим эталоном.
В большинстве современных статей проблема связи генотипа с фенотипом формулируется в терминах карт генотипа и фенотипа. Первая карта GP была представлена Ричардом Левонтином в его книге «Генетическая основа эволюционных изменений» (Lewontin, 1974a; , рис. ).Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разделен на четыре этапа: (1) средний фенотип является производным от развития отдельных генотипов в различных средах; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, у которых будет потомство; (3) личность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутация и рекомбинация, изменяют положение в G-пространстве.
В другом распространенном графическом представлении ( Рисунок ) точка в пространстве G и соответствующая ей точка в пространстве P соответствуют генотипу и фенотипу отдельного человека (Fontana, 2002; Landry and Rifkin, 2012). При таком представлении абстрактный объект, который мы определили выше как отношение GP, будет соответствовать «движению» в пространстве генотипа, связанному с «движением» в пространстве фенотипа (или, лучше, сумме нескольких «ходов» в генотипе, и пространства фенотипов, потому что несколько различных геномов могут нести два альтернативных аллеля данного отношения GP).В третьем представлении, выдвинутом Вагнером (1996; Рисунок ), отдельные гены связаны с отдельными признаками.
Хотя эти три графических представления карт GP могут облегчить наше понимание определенных аспектов биологии, во всех из них связь GP и дифференциальная точка зрения непросто понять. Вызывает недоумение то, что первым, кто нарисовал такую карту GP, был Ричард Левонтин, красноречивый сторонник дифференциальной точки зрения (см., Например, его предисловие к Oyama, 2000, шедевр убеждения).Поскольку эти графики сосредоточены на отдельных, а не на различных объектах, мы считаем, что эти три представления неявно побуждают нас вернуться к более интуитивному представлению об одном генотипе, связанном с одним фенотипом. Упущение из виду дифференциальной точки зрения может также происходить с точки зрения молекулярной биологии, где белки рассматриваются как имеющие причинные эффекты сами по себе, такие как фосфорилирование субстрата или связывание с последовательностью ДНК. Из-за двух запутанных определений гена, кодирующего белок или вызывающего фенотипические изменения (Griffiths and Stotz, 2013), легко перейти от дифференциальной точки зрения к недифференциальной точке зрения на отношения GP.
Таким образом, многие современные ментальные представления о связи между генотипом и фенотипом неявно отвергают дифференциальную точку зрения. Теперь мы покажем, что дифференциальная точка зрения совместима с тем фактом, что на фенотипические черты влияет сложная комбинация множества факторов, и что мы можем найти соответствующее схематическое представление отношений GP.
Проблема плейотропии
Разложение организма на элементарные единицы, такие как анатомические структуры, сыграло важную роль во многих биологических дисциплинах, таких как физиология, палеонтология и эволюция.Однако проблема состоит в том, чтобы определить декомпозицию на символы, наиболее подходящую для интересующего вопроса. Для вопросов, связанных с взаимоотношениями между органами разных людей или видов (например, гомология), может быть целесообразно сохранить традиционную декомпозицию на анатомические структуры (Wagner, 2014). Ричард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют квазинезависимые единицы эволюционных изменений (Левонтин, 1978; Вагнер, 2000).Их определение касается абсолютных черт, наблюдаемых у отдельных организмов (например, формы крыла или количества пальцев у особи), и, таким образом, далеко от дифференциальной точки зрения. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые атрибуты организма на множество элементарных вариаций GP, которые накапливались в течение нескольких поколений, начиная с начального состояния. Мы настаиваем на том, что с этой точки зрения персонажи — это не конкретные объекты (например, кожа), а абстрактные сущности, определяемые существованием различий между двумя возможными наблюдаемыми состояниями (например, цветом кожи).В качестве аналогии можно представить два способа изготовления поношенной кожаной обуви определенной формы. Можно либо собрать разные атомы в одну и ту же организацию, либо можно купить обувь в магазине и затем подвергнуть ее воздействию ряда механических сил. Мы естественно склонны сравнивать организмы с машинами и мыслить категориями частей, которые должны быть собраны, чтобы образовать функциональное целое. Однако безудержная метафора дизайнера или производителя неадекватна для понимания происхождения современных организмов (Coen, 2012).Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые произошли последовательно в течение эволюционного времени, а не во время развития. Исходное состояние является гипотетическим предком исследуемого организма.
Наблюдается, что некоторые мутации (квалифицируемые как плейотропные) затрагивают сразу несколько органов, в то время как другие изменяются только по одному (Paaby and Rockman, 2013; Zhang and Wagner, 2013). Для плейотропных мутаций мы считаем, что отношения GP должны включать все фенотипические изменения (в разных органах, на разных стадиях и т. Д.)) связано с генетическим различием. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменениями плотности потовых и молочных желез в азиатских популяциях (Kamberov et al., 2013). В таких случаях отношения GP являются взаимно-множественными. Рассмотрение кожи и глаз как независимых анатомических модулей человеческого тела может показаться подходящим для многих эволюционных изменений, но это несколько неадекватно в тех случаях, когда эти два органа развили новую черту пигментации одновременно через единственную мутацию в гене SLC45A2 (Лю и другие., 2013). Рассуждения в терминах отношений GP снимают проблему нахождения релевантной декомпозиции на элементарные анатомические структуры. Сами элементарные отношения GP выглядят как адекватные полунезависимые модули, комбинация которых может объяснить наблюдаемые характеристики организма.
Проблема непрерывных сложных признаков
Согласно дифференциальной концепции отношений GP, одним из важнейших моментов является разложение наблюдаемых признаков на серию полунезависимых фенотипических вариаций, то есть выявление элементарных изменений, произошедших в ходе эволюции.Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разными формами листьев дает потомство, которое демонстрирует составной диапазон промежуточных форм листьев. Анализ главных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и которые, по-видимому, вызваны разными геномными регионами (Langlade et al., 2005). Это в какой-то мере говорит о том, что «сумма затемняет части.«То, что мы традиционно считаем сложными, можно сделать из более простых, более поддающихся генетическому анализу. Другой яркий пример — пигментация брюшной полости в группе Drosophila dunni . Взятые как единственная переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение паттернов взрослых на анатомические субъединицы нарушает дискретный генетический контроль для каждого суб-признака (Hollocher et al., 2000). Более известный случай — это эволюция цвета тела у пляжных мышей.Разница в цвете между светлыми пляжными мышами и темными мышами может быть разложена на отдельные фенотипы (спинной оттенок, дорсальная яркость, ширина полосы на хвосте и дорсовентральная граница), которые все связаны с различными мутациями в гене Agouti ( Linnen et al., 2013; Рисунок ). Кажется, что каждый генетический локус Agouti предназначен для спецификации пигментации в данной части тела. Вместе они образуют группу тесно связанных локусов, которые связаны с изменениями пигментации шерсти.
Эволюция светлых пляжных мышей вызвана несколькими мутациями с отчетливыми эффектами пигментации в локусе Agouti . Темный и светлый фенотипы можно разложить на четыре фенотипических признака, которые связаны с различными однонуклеотидными полиморфизмами (SNP, цветные точки), расположенными в гене Agouti . Показаны только SNP с предполагаемым коэффициентом отбора легкого аллеля выше 0,1. Кодирующие экзоны представлены темными прямоугольниками, а непереведенные экзоны — белыми.По материалам Linnen et al. (2013).
В то время как сложные черты не всегда могут быть сведены к набору простых взаимоотношений GP, возможно, что такие черты, как рост взрослого человека, наиболее символический количественный признак, который, по прогнозам, состоит из множества генетических эффектов небольшого размера (Fisher, 1930), также могут быть разложены на элементарные вариации, каждая из которых объясняет более дискретные суб-черты. В то время как некоторые детерминанты человеческого роста, такие как LIN28B , были связаны с ростом взрослого человека в разном возрасте, другие гены достигли статистической значимости только в исследованиях, посвященных конкретным стадиям, с упором на рост плода и скорость роста в период полового созревания (Lettre, 2011).Другими словами, эти данные предполагают, что рост человека может быть составной чертой, которая модулируется несколькими отношениями GP, каждая из которых действует на разных фазах развития.
Проблема эпистаза и GxE
Взаимодействие между генами и средой (GxE) происходит, когда фенотипический эффект данного генетического изменения зависит от параметров окружающей среды. Сходным образом эпистаз или взаимодействие GxG происходит, когда фенотипический эффект данного генетического изменения зависит от аллельного состояния по крайней мере одного другого локуса (Phillips, 2008; Hansen, 2013).Появляется все больше доказательств того, что взаимодействия GxG и GxE имеют фундаментальное значение для понимания эволюции и наследования сложных признаков (Gilbert and Epel, 2009; Hansen, 2013). Мы предполагаем, что оба явления могут быть интегрированы в базовую дифференциальную структуру GP, где взаимодействия GxG и GxE создают слой зависимости от контекста и приводят к различиям, заключенным в различиях.
Разница в цветовой пигментации между темными и светлыми пляжными мышами, упомянутая ранее ( рис. ), связана не только с мутациями в Agouti , но и с кодирующей мутацией в гене MC1R , которая уменьшает пигментацию (Steiner et al. al., 2007; Рисунок ). Эффект мутации MC1R виден только в присутствии связанного со светом производного гаплотипа Agouti . Здесь локус Mc1R считается эпистатически взаимодействующим с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два возможных фенотипических различий (изменение пигментации шерсти или отсутствие изменений вообще). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь в локусе Agouti ).Таким образом, дифференциальная точка зрения остается относительно простой для двухлокусных взаимодействий: контекстная зависимость фенотипа транслируется в выбор между двумя возможными фенотипическими различиями. Мы предлагаем, чтобы отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, включали все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах. Среди всех возможных фенотипических вариаций наблюдаемые фенотипические различия определяются другими генетическими локусами.В общем, взаимодействия GxG включают несколько сайтов, которые рассредоточены по геному (Bloom et al., 2013).
Взаимодействие генов за средой (GxE) и GxG. (A) Кодирующая мутация npr-1 влияет на агрегацию нематод при 21% уровне кислорода, но не при 10% (Andersen et al., 2014). (B) Кодирующая мутация Mc1R влияет на пигментацию тела мыши в присутствии доминантных светлых аллелей Agouti , но не на гомозиготном фоне Agouti по рецессивному темному аллелю (Steiner et al., 2007).
Примером взаимодействия GxE (см. Также Рисунок ) является встречающийся в природе аллель потери функции brx у растений Arabidopsis , который связан с ускоренным ростом и повышенной приспособляемостью в кислых почвах, а также с сильным снижение роста корней по сравнению с диким типом в нормальных почвах (Gujas et al., 2012). Взаимодействия GxE обычно анализируются в форме нормы реакции , которая представляет все наблюдаемые черты единственного генотипа в различных средах (Johannsen, 1911; Sarkar, 1999).В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Связанное с этим фенотипическое изменение, таким образом, представляет собой разницу между двумя нормами реакции. Примером из учебника является правило изменения температуры и размера в C. elegans . Как и большинство других животных, нематоды C. elegans и растут крупнее при низкой температуре, но лабораторный штамм дикого типа с температурой C.elegans , происходящих из Гавайев, не показывает изменений в размере тела при различных температурах. Аминокислотная замена в кальций-связывающем белке ответственна за снижение способности гавайского штамма расти при низкой температуре (Kammenga et al., 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанная с ней взаимосвязь GP включает разницу между этими двумя наклонами.
Диапазон фенотипических вариаций, воплощенных в отношениях GP, подверженных взаимодействиям GxG и GxE, может быть довольно огромным, особенно в случаях, когда несколько тканей затронуты одной и той же мутацией, и когда на фенотипические вариации каждой ткани влияют другие геномные локусы и по условиям окружающей среды.Фактически, фенотипические эффекты мутации всегда зависят от других фрагментов ДНК из того же генома, так что любые отношения GP могут рассматриваться как переживание эпистаза. Другими словами, генетический локус, влияющий на фенотип, никогда не действует независимо от других последовательностей ДНК. Например, данный аллель опсина будет приводить к определенным свойствам цветового зрения только в том случае, если глаз сформирован и если этот глаз получает свет во время своего развития, что позволяет формировать эффективные нейронные цепи зрения. Чтобы дифференцированный взгляд был податливым, мы советуем не учитывать все возможные генетические фоны и условия окружающей среды, а ограничить возможности потенциальной средой и сегрегацией аллелей, относящихся к интересующей популяции (Sober, 1988).
Таким образом, при наличии эпистаза или взаимодействий GxE генетическое изменение связано не с одним фенотипическим различием, а с множеством возможных фенотипических различий, одно из которых будет достигнуто в зависимости от окружающей среды и генетического фона. Зависимость от контекста может быть схематично представлена как различия GP, встроенные в различия других генотипов и окружающей среды.
Дифференциальный взгляд на генетические и экологические эффекты на фенотипы
Как подчеркивалось многими авторами (в первую очередь Waddington, 1957; Oyama, 2000; Keller, 2010), гены и окружающая среда действуют совместно на фенотип, и в большинстве случаев это так. невозможно отделить эффект одного от другого.Здесь мы показываем, что рассуждения о различиях помогают прояснить сравнение между генетическим и средовым влиянием на фенотипы. Однако мы выявляем определенные случаи, когда сравнение остается затруднительным.
По аналогии с отношениями GP, мы можем определить отношения между средой и фенотипом как средовую вариацию (две среды), связанные с ней фенотипические изменения (различные фенотипические состояния) и их отношения. Например, у многих видов черепах изменение температуры во время развития яйца связано с различием пола самец / самка ( Рисунок ), и по крайней мере шесть переходов от экологического к генетическому определению пола ( Рисунок ) произошли в филогенезе черепах. (Pokorná, Kratochvíl, 2009).В этом случае можно сравнить экологические и генетические эффекты: половые хромосомы и температура имеют одинаковый фенотипический эффект на черепах. Такие наблюдения привели Вест-Эберхарда (2003, 2005) к предложению гипотезы «гены как последователи», которая предполагает, что новые фенотипические состояния с большей вероятностью возникают сначала из-за изменения окружающей среды, чем из-за генетической мутации, и что мутации происходят только позже, при изменении порога проявления новой черты. Вест-Эберхард (2003, 2005) экстраполировал от различий, сегрегированных внутри популяций (различие № 2), до различий, которые возникли во времени во время эволюции популяции (различие № 1).
Отношения окружающая среда – фенотип и отношения GP для определения пола у черепах. (A) У некоторых видов температура во время эмбрионального развития определяет пол взрослой особи. (B) У других пол определяется половыми хромосомами.
Независимая эволюция направленной лево-правой асимметрии от симметричных предков в нескольких ветвях предоставила главный аргумент в пользу гипотезы «ген как последователь» (Palmer, 2004). В соответствии с этой структурой оговаривается, что направленная асимметрия, когда все индивиды односторонние, часто эволюционировала из состояния «случайной асимметрии», где направленность будет зависеть от факторов окружающей среды и, таким образом, будет варьироваться между генетически идентичными индивидами.Например, самая сильная клешня омара будет развиваться в зависимости от использования и имеет априори равную вероятность развития левой или правой стороны. Мы можем видеть, как здесь применяется формула «гены как последователи»: окружающая среда вызывает асимметрию, и позже в эволюции некоторые генетические эффекты могут изменить ее направленность в ту или иную сторону. Но в то время как асимметрия «возникает до того, как существует генетическая изменчивость, которая ее контролирует», дифференциальная точка зрения проясняет, что генетическое влияние на направленность несопоставимо с влиянием окружающей среды, которое запускает асимметрию.Генетическое изменение приводит к переключению между окончательным 100% односторонним состоянием и начальным состоянием, при котором 50% случаев являются правосторонними и 50% левосторонними. Напротив, два альтернативных фенотипических состояния, возникающих в результате изменения окружающей среды, считаются 100% правыми и 100% левыми. Этот пример показывает, что для точности важно явно указать различия, которые учитываются в отношениях GP.
Дифференциальное представление обеспечивает теоретическую основу, которая может помочь в разработке экспериментов для исследования правильных переменных: можно сравнивать разные генотипы в фиксированной среде (классические отношения GP), сравнивать реакцию фиксированного генотипа на две разные среды (фенотипическая пластичность ) или сравнить чувствительность двух разных генотипов к двум различным средам (при этом фенотипическая вариация становится различием при различии ; см., например, Engelman et al., 2009; Томас, 2010).
Были разработаны различные количественные методы, позволяющие отделить генетические эффекты от воздействия окружающей среды и количественно оценить взаимодействия GxE (Lynch and Walsh, 1998). Тем не менее, в определенных ситуациях может оказаться невозможным биологически значимым образом отделить генетическое воздействие от воздействия окружающей среды, даже если рассуждать с точки зрения различий (Lewontin, 1974b). Популяции жука Calathus melanocephalus включают две морфы: длиннокрылые и короткокрылые (Schwander, Leimar, 2011).Длиннокрылые морфы развиваются только от гомозиготных особей по рецессивному аллелю, сегрегационному в популяции, и только при хороших условиях питания. В этом случае генетические эффекты и эффекты окружающей среды переплетаются (фигуры , фигуры ). В теоретическом случае популяции, состоящей только из короткокрылых гетерозиготных животных, которые были выращены в условиях голодания и длиннокрылых, и гены, и среда несут ответственность за различие крыльев между особями, и невозможно оценить долю окружающей среды и генетические эффекты, потому что гены и окружающая среда действуют на разных уровнях сложной причинной связи между генотипами и фенотипами.
Взаимосвязь окружающая среда-фенотип и перспективы взаимосвязи GP для полиморфизма длины крыла у жука Calathus melanocephalus . (A) С точки зрения взаимосвязи окружающая среда-фенотип, изменение условий питания связано с изменением размера крыла, но только с гомозиготным фоном для рецессивного аллеля (-1 ) локуса размера крыла. (B) С точки зрения взаимоотношений GP, генетическое изменение в локусе размера крыла связано с изменением размера крыла, но только в хороших условиях питания.
Другой случай, который ставит под сомнение классическое различие между окружающей средой и генетикой, — это когда добавление определенных симбиотических бактерий изменяет фенотип хозяина. Мыши, которых кормили штаммом бактерий Lactobacillus , демонстрируют сниженное поведение, связанное с тревогой, по сравнению с контрольными мышами, которых кормили бульоном без бактерий (Bravo et al., 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного симбионта кишечника. Причиной фенотипических различий является не простое изменение последовательности ДНК или простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между присутствием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — которые содержат ДНК, мутации которой также могут изменять фенотип хозяина.
В заключение, рассуждения о различиях могут помочь прояснить сравнение между генетическим воздействием и влиянием окружающей среды на фенотипы. Однако проблемы здесь очень простые. Поскольку гены и окружающая среда действуют на разных уровнях сложной причинной связи между генотипами и фенотипами, в некоторых случаях невозможно разделить обе причины.
A Разъяснение терминологии Прибыль / потеря и разрешающая / поучительная
Фенотипические различия, по-видимому, подпадают под две основные категории: либо наличие / отсутствие чего-либо (например, волос на теле или способность переваривать молоко), либо переключение между две альтернативы, которые присутствуют обе (например, два цвета волос).Точно так же на стороне генотипа мутация может соответствовать наличию / отсутствию соответствующей последовательности ДНК или полиморфизму нуклеотидов. Дифференциальная перспектива делает очевидным, что потеря фенотипа не обязательно связана с потерей генетического материала, и наоборот. Например, эволюционное увеличение темных пигментов, покрывающих всю шерсть животных, часто было связано с потерей гена Mc1R (Gompel and Prud’homme, 2009). Более того, как один из нас ранее отмечал (приложение Stern and Orgogozo, 2008), выигрыш или потеря для фенотипа субъективны.Например, выпадение волос можно рассматривать как прирост голого эпидермиса. Большинство эпидермальных клеток насекомых дифференцируются в одно из этих двух альтернативных состояний, и в обоих состояниях задействованы большие сети регуляции генов. Неясно, какое фенотипическое состояние представляет собой выигрыш или потерю по сравнению с другим. Даже на генотипической стороне определение потерь и прибылей может быть трудным. Вставка мобильного элемента может сбивать ген, тогда как делеция может иногда создавать новый сайт связывания для активатора транскрипции.Фактически, эволюционный прирост экспрессии desatF у D. melanogaster происходит посредством серии из трех делеций, каждая из которых создает мотив гексамера, который необходим для экспрессии desatF (Shirangi et al., 2009).
Точно так же дифференциальный взгляд на воздействие окружающей среды подчеркивает ошибочность различия между разрешающими и поучительными сигналами. Пермиссивный сигнал связан с наличием / отсутствием фенотипа, а инструктивный сигнал — со сдвигом между двумя присутствующими альтернативами.Как указывалось выше, эти различия на фенотипическом уровне нечеткие.
В заключение мы предлагаем с осторожностью использовать терминологию «прибыль / убыток» и поучительную / разрешительную терминологию.
Таксономически устойчивые отношения GP
Ожидается, что мутация вызовет в некоторой степени воспроизводимые фенотипические вариации в популяции. Такая воспроизводимость фенотипических результатов необходима для генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998).В самом деле, новообразованный аллель, который будет генерировать еще один фенотип каждый раз, когда он окажется в другом организме, не будет подвергнут естественному отбору. Рассуждения с точки зрения вариации, а не рассмотрение аллелей как изолированных объектов, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор действует непосредственно на аллельную изменчивость, которая последовательно связана с данной фенотипической изменчивостью, которая сама по себе является отношением GP. Таким образом, отношения GP являются базовой единицей эволюционных изменений, на которую действует естественный отбор.
Важным открытием последних 20 лет является то, что вариации в определенных генетических локусах вызывают сравнимые фенотипические вариации не только у разных особей одной популяции, но и у чрезвычайно разных таксонов (Martin and Orgogozo, 2013b). Другими словами, определенные отношения GP устойчивы с таксономической точки зрения и присутствуют в большом диапазоне видов. Это означает, что генетический и экологический фон оставался относительно постоянным или появлялся неоднократно на протяжении эволюции, чтобы позволить генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах.Об этом важном открытии почти 50 лет назад никто не подозревал. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сопоставимую сингулярность на генетическом уровне, включая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 1963). Как однажды предложил Майр (1963) в 1963 году: «Многое из того, что было изучено о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» так же верна как в эволюции, так и в повседневной жизни »(Mayr, 1963).Другими словами, генетические локусы, которые делают человека мужчиной, должны были отличаться от локусов, которые делают собаку или рыбу. Позже, в 80–90-х годах, несколько исследователей предположили совершенно противоположное, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 1978; Perutz, 1983; Stewart et al., 1987; Carroll et al. ., 2005), но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). На сегодняшний день накопление данных о мутациях, ответственных за естественную изменчивость, ясно показывает, что разнообразие живых организмов имеет общую генетическую основу как минимум по трем пунктам.Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с сохраненными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых генетическая основа была хотя бы частично идентифицирована (154 случая из 160), вызваны мутациями в гомологичных генах, и очень немногие (6/160) связаны с новыми генами, которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b).В-третьих, было показано, что множественные случаи сходных фенотипических изменений связаны с мутациями одних и тех же гомологичных генов в независимых линиях ( Таблица ), иногда на больших филогенетических расстояниях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-переносчика SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментированными глазами, кожей , волосы и перья у людей и кур (Xu et al., 2013; Рисунок ). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад — моллюска, змеи и иглобрюха — через одну и ту же аминокислотную замену в консервативном гене (Bricelj et al., 2005 ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения сейчас обнаружены для более чем 100 генов у животных и растений (Martin and Orgogozo, 2013b). Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере определенных фенотипических различий существует относительно мало генетических путей. ведут в Рим (Stern, 2013).В настоящее время не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев у одного вида бабочек Heliconius , обеспечивает аналогичные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой, а человека мужчиной, теперь частично объясняется уникальным набором таксономически устойчивых отношений GP, которые обнаруживаются в нескольких ветвях родословной.
Определенные отношения между средой и фенотипом также устойчивы с таксономической точки зрения.Например, у большинства таксонов размер тела зависит от питания; дефицит железа может вызвать анемию, а некоторые токсичные соединения могут быть смертельными. У эктотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, являются ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип более распространенными. Кроме того, является предметом споров, представляют ли таксономически устойчивые отношения GP исключительную и небольшую часть или значительную долю всех отношений GP.В любом случае существование таксономически устойчивых взаимоотношений с терапевтами теперь очевидно и должно быть широко принято исследовательским сообществом.
Некоторые из наиболее ярких учений современной биологии включают открытие того, что живые существа имеют один и тот же генетический материал (ДНК или РНК), один и тот же генетический код (за некоторыми исключениями) и один и тот же базовый клеточный механизм. Таким образом, совсем не парадоксально, что индивидуальные различия строятся на сходствах, и открытие того, что определенные отношения GP сохраняются в течение длительного времени эволюции, дополняет картину.
Точная предсказательная сила, являющаяся результатом существования таксономически устойчивых взаимосвязей GP, редко встречается в биологии и только начинает использоваться в полной мере. Сохранение связей GP на больших расстояниях теперь полностью оправдывает использование подходов сравнительной генетики для решения прагматических проблем. Например, одомашнивание сельскохозяйственных культур принимало форму аналогичного давления отбора у многих видов, и теперь у нас есть экспериментальные доказательства того, что этот процесс неоднократно включал мутации в одном и том же наборе консервативных генов (Paterson et al., 1995; Мартин и Оргогозо, 2013b). Это наблюдение открывает интересные приложения, поскольку мы можем использовать этот появляющийся массив генетических знаний для помощи в одомашнивании будущих культур или использовать стратегии с использованием маркеров для создания и поддержания биоразнообразия сельскохозяйственных культур (Lenser and Theißen, 2013). Предсказуемость GP уже используется при идентификации штаммов, у которых развилась устойчивость к различным стратегиям борьбы с вредителями, причем в крайних случаях нацелена на устойчивость к противомалярийным препаратам у паразитов Plasmodium (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более важно, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статуса вредителей (Ffrench-Constant et al., 2004; Martin, Orgogozo, 2013b).
Кроме того, повторяемость генетической основы фенотипической изменчивости предполагает, что клинические исследования также могут выиграть от генетических исследований большого диапазона модельных видов (Robinson and Webber, 2014).Например, естественные вариации толерантности к метотрексату, химиотерапевтическому препарату, были сопоставлены у плодовых мух Drosophila с генами, чьи человеческие ортологи также связаны с реакцией пациентов на этот препарат (Кислухин и др., 2013), таким образом увеличивая использование модельных организмов в качестве моделей болезней.
К классификации фенотипов на основе генов
Одним из оригинальных аспектов определения связи GP с точки зрения индивидуальных взаимоотношений GP является то, что он позволяет классифицировать фенотипы в соответствии с их лежащей в основе генетической основой.На первом уровне взаимосвязи GP, затрагивающие разные области одного и того же гена и приводящие к сопоставимым фенотипическим результатам, могут быть сгруппированы вместе. Простые случаи взаимодействий GxG были обнаружены между прочно сцепленными мутациями, как правило, внутри кодирующей последовательности или внутри регуляторного элемента cis-, когда они вызывают неаддитивный эффект на фенотип. Например, наблюдали, что конкретная мутация в энхансере вызывает различные сдвиги в паттерне экспрессии нижележащего кодирующего гена в зависимости от соседней последовательности ДНК (Frankel et al., 2011; Роджерс и др., 2013). Аналогичным образом, аминокислотные мутации в гене гемоглобина, как было обнаружено, увеличивают или уменьшают сродство к кислороду в зависимости от аллельного состояния других сайтов (Natarajan et al., 2013). В таких случаях интуитивно легко сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же фенотипический признак.
Отсутствие пигментов меланина у животных было связано с мутациями в нескольких генах, включая OCA2 , kit-лиганд или Mc1R (обзор в Gompel and Prud’homme, 2009; Liu et al., 2013). Принимая во внимание, что отсутствие меланина традиционно считается одним из характерных состояний, альбинизмом, независимо от лежащей в основе генетической основы, мы предлагаем здесь различать альбинизм, связанный с OCA2 , от альбинизма, ассоциированного с Mc1R , или от альбинизма, ассоциированного с любым другим геном. Один интерес в разложении вариаций в живом мире на эти множественные элементарные отношения GP состоит в том, что эти элементы затем могут быть сгруппированы вместе в последовательно увеличивающиеся группы.Элементарные фенотипические изменения с участием разных генов, которые являются частью одного и того же генетического пути, также могут быть сгруппированы вместе как сопутствующие компоненты одного и того же механизма модуляции фенотипа. Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9 и рецептору TGF-β BMPR1B, которые все неоднократно были связаны с вариациями функции яичников у людей и домашних пород овец (обзор в Luong et al., 2011).
Еще одно важное следствие перспективы взаимоотношений GP состоит в том, что очевидно различные фенотипические изменения, вызванные сходными генетическими локусами у разных организмов, могут быть исследованы дополнительно, чтобы выявить то, что может быть общим основным фенотипическим изменением (Deans et al., 2015). Например, личинки мух и нематодные черви имеют различное поведение при поиске пищи, но мутации в одном и том же ортологическом гене ( для / egl-4 ), как было показано, изменяют интенсивность поведения поиска пищи у обоих организмов (Osborne et al. , 1997; Mery et al., 2007; Hong et al., 2008). Таким образом, вероятно, что базовое изменение поведения, лежащее в основе, казалось бы, отличительных изменений в поиске пищи мухами и нематодами, представляет собой сохраненные отношения GP между нематодами и мухами.Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к болезни человека? Можно ли считать, что фенотип мыши похож на состояние человека, если его генетическая основа иная? Мы и другие предсказываем, что поиск ортологичных фенотипов или «фенологов» (McGary et al., 2010) станет важной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
Заключение
В этой статье мы возвращаем дифференциальную концепцию гена (Schwartz, 2000) в нашу структуру для понимания карты GP. Дифференциальный взгляд на отношения GP помогает прояснить генетическое и экологическое влияние на фенотипы и их связь. Это также открывает новые пути мышления, в частности, в отношении декомпозиции наблюдаемых функций внутри организма и представления карт GP. Кроме того, наличие таксономически устойчивых взаимоотношений с GP побуждает беззастенчиво использовать сравнительную генетику для прогнозирования генетической основы фенотипической изменчивости в различных группах организмов, и эта прогностическая сила имеет важный потенциал для трансляционных исследований в агрономии и клинических исследованиях.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы глубоко признательны Мари-Анн Феликс за полезные обсуждения и Томасу Прадо за то, что привлекли наше внимание к нескольким актуальным статьям. Мы также благодарим Джузеппе Бальдаччи, Мари-Анн Феликс, Пьера-Анри Гуйона, Александра Пелуффо, Марка Сигала, Дэвида Стерна и рецензентов за их проницательные комментарии к рукописи.Исследование, приведшее к этой статье, финансировалось Европейским исследовательским советом в рамках Седьмой рамочной программы Европейского сообщества (грантовое соглашение FP7 / 2007-2013 № 337579) и Фондом Джона Темплтона (грант № 43903).
Ссылки
- Андерсен Э. К., Блум Дж. С., Герке Дж. П., Кругляк Л. (2014). Вариант нейропептидного рецептора npr-1 является основным детерминантом роста и физиологии Caenorhabditis elegans . PLoS Genet. 10: e1004156 10.1371 / journal.pgen.1004156 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Армстронг М. Д., Тайлер Ф. Х. (1955). Исследования фенилкетонурии. I. Ограничение приема фенилаланина при фенилкетонурии. J. Clin. Вкладывать деньги. 34 565–580. 10.1172 / JCI103105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Bloom J. S., Ehrenreich I. M., Loo W. T., Lite T.-L. В., Кругляк Л. (2013). Выявление источников потери наследуемости при скрещивании дрожжей. Природа 494 234–237. 10.1038 / nature11867 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Бойкот А. Э., Дайвер К., Гарстанг С. Л., Тернер Ф. М. (1931). Наследование левосторонности у limnæa peregra (Mollusca, Pulmonata). Philos. Пер. R. Soc. Лондон. Сер. B 219 51–131. 10.1098 / rstb.1931.0002 [CrossRef] [Google Scholar]
- Браво Дж. А., Форсайт П., Чу М. В., Эскаравадж Э., Савиньяк Х. М., Динан Т. Г. и др. (2011). Проглатывание штамма Lactobacillus регулирует эмоциональное поведение и экспрессию центрального рецептора ГАМК у мыши через блуждающий нерв. Proc. Natl. Акад. Sci. США 108 16050–16055. 10.1073 / pnas.1102999108 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Бричелж В. М., Коннелл Л., Коноки К., Маккуори С. П., Шойер Т., Кэттералл В. А. и др. (2005). Мутация натриевых каналов, приводящая к устойчивости к сакситоксину у моллюсков, увеличивает риск PSP. Природа 434 763–767. 10.1038 / nature03415 [PubMed] [CrossRef] [Google Scholar]
- Кэрролл С. Б., Гренье Дж., Уэтерби С. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных .Молден, Массачусетс: John Wiley & Sons. [Google Scholar]
- Кассирер Э. (1910). Substance et Fonction 2010 Эдн Париж: Nabu Press. [Google Scholar]
- Коэн Э. (2012). От клеток к цивилизациям: принципы изменения, формирующие жизнь . Принстон, Нью-Джерси: Издательство Принстонского университета; 10.1515 / 9781400841653 [CrossRef] [Google Scholar]
- Колозимо П. Ф., Хосеманн К. Э., Балабхадра С., Вильярреал Г. младший, Диксон М., Гримвуд Дж. И др. (2005). Распространенная параллельная эволюция колюшек путем многократной фиксации аллелей эктодисплазина. Наука 307 1928–1933. 10.1126 / science.1107239 [PubMed] [CrossRef] [Google Scholar]
- Докинз Р. (1982). Расширенный фенотип: долгая досягаемость гена . Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- Deans A. R., Lewis S. E., Huala E., Anzaldo S. S., Ashburner M., Balhoff J. P. и др. (2015). Пробираемся сквозь фенотипы. PLoS Biol. 13: e1002033 10.1371 / journal.pbio.1002033 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Данбар Х.Э., Уилсон А. С., Фергюсон Н. Р., Моран Н. А. (2007). Термостойкость тли определяется точечной мутацией у бактериальных симбионтов. PLoS Biol. 5: e96 10.1371 / journal.pbio.0050096 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Энгельман К. Д., Баурли Дж. У., Чиу Й.-Ф., Жубер Б. Р., Левингер Дж. П., Меннер М. Дж. И др. (2009). Обнаружение взаимодействий между геном и окружающей средой в данных ассоциации по всему геному. Genet. Эпидемиол. 33 (Дополнение 1) S68 – S73. 10.1002 / gepi.20475 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Falk R. (1984). Ген в поисках идентичности. Hum. Genet. 68 195–204. 10.1007 / BF00418388 [PubMed] [CrossRef] [Google Scholar]
- Фельдман К. Р., Броди Д., младший, Броди Д., III, Пфрендер М. Э. (2012). Ограничение формирует схождение тетродотоксин-устойчивых натриевых каналов змей. Proc. Natl. Акад. Sci. США 109 4556–4561. 10.1073 / pnas.1113468109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ffrench-Constant R.Х., Даборн П. Дж., Ле Гофф Г. (2004). Генетика и геномика устойчивости к инсектицидам. Trends Genet. 20 163–170. 10.1016 / j.tig.2004.01.003 [PubMed] [CrossRef] [Google Scholar]
- Фишбах М. А. (2009). Антибиотики от микробов: убивают. Curr. Opin. Microbiol. 12 520–527. 10.1016 / j.mib.2009.07.002 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Фишер Р. А. (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон; 10.5962 / bhl.title.27468 [CrossRef] [Google Scholar]
- Fontana W. (2002). Моделирование «эво-дево» с помощью РНК. BioEssays News Rev. Mol. Клетка. Dev. Биол. 24 1164–1177. 10.1002 / bies.10190 [PubMed] [CrossRef] [Google Scholar]
- Франкель Н., Эрезилмаз Д. Ф., МакГрегор А. П., Ван С., Пайре Ф., Стерн Д. Л. (2011). Морфологическая эволюция, вызванная множеством малозаметных замен в регуляторной ДНК. Природа 474 598–603. 10.1038 / nature10200 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Geffeney S.Л., Фудзимото Э., Броди Э. Д., III, Броди Э. Д., младший, Рубен П. К. (2005). Эволюционная диверсификация ТТХ-резистентных натриевых каналов во взаимодействии хищник-жертва. Природа 434 759–763. 10.1038 / nature03444 [PubMed] [CrossRef] [Google Scholar]
- Gerstein M. B., Bruce C., Rozowsky J. S., Zheng D., Du J., Korbel J. O., et al. (2007). Что такое ген пост-ENCODE? История и обновленное определение. Genome Res. 17 669–681. 10.1101 / gr.6339607 [PubMed] [CrossRef] [Google Scholar]
- Gilbert S.Ф. (2000). «Гены классические и гены развития: различные варианты использования генов в эволюционном синтезе», в Концепция гена в развитии и эволюции редакторы Beurton P., Falk R., Rheinberger H.-J. (Кембридж, Нью-Йорк: издательство Кембриджского университета;). [Google Scholar]
- Гилберт С. Ф., Эпель Д. (2009). Экологическая биология развития: интеграция эпигенетики, медицины и эволюции . Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
- Gjuvsland A.Б., Вик Дж. О., Борода Д. А., Хантер П. Дж., Омхольт С. В. (2013). Преодоление разрыва между генотипом и фенотипом: что для этого нужно? J. Physiol. 591 2055–2066. 10.1113 / jphysiol.2012.248864 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Gompel N., Prud’homme B. (2009). Причины повторной генетической эволюции. Dev. Биол. 332 36–47. 10.1016 / j.ydbio.2009.04.040 [PubMed] [CrossRef] [Google Scholar]
- Граур Д., Чжэн Ю., Азеведо Р. Б. Р. (2015). Эволюционная классификация геномной функции. Genome Biol. Evol. 7 642–645. 10.1093 / gbe / evv021 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Griffiths P., Stotz K. (2013). Генетика и философия: введение . Кембридж: Издательство Кембриджского университета; 10.1017 / CBO9780511744082 [CrossRef] [Google Scholar]
- Гуджас Б., Алонсо-Бланко К., Хардтке С. С. (2012). Природные аллели потери функции Arabidopsis brx придают корням адаптацию к кислой почве. Curr. Биол. 22 1962–1968 гг.10.1016 / j.cub.2012.08.026 [PubMed] [CrossRef] [Google Scholar]
- Хансен Т. Ф. (2013). Почему эпистаз важен для отбора и адаптации. Evolution 67 3501–3511. 10.1111 / evo.12214 [PubMed] [CrossRef] [Google Scholar]
- Холлохер Х., Хэтчер Дж. Л., Дайресон Э. Г. (2000). Генетический анализ и анализ развития различий пигментации брюшной полости у разных видов в подгруппе Drosophila dunni . Evol. Int. J. Org. Evol. 54 2057–2071. 10.1111 / j.0014-3820.2000.tb01249.x [PubMed] [CrossRef] [Google Scholar]
- Хонг Р. Л., Витте Х., Соммер Р. Дж. (2008). Природные вариации в привлечении феромонов насекомых Pristionchus pacificus связаны с протеинкиназой EGL-4. Proc. Natl. Акад. Sci. США 105 7779–7784. 10.1073 / pnas.0708406105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Хоул Д., Говиндараджу Д. Р., Омхольт С. (2010). Феномика: следующий вызов. Nat. Преподобный Жене. 11 855–866.10.1038 / nrg2897 [PubMed] [CrossRef] [Google Scholar]
- Johannsen W. (1911). Генотипическая концепция наследственности. г. Nat. 45 129–159. 10.1086 / 279202 [CrossRef] [Google Scholar]
- Джонс Ф. К., Грабхер М. Г., Чан Ю. Ф., Рассел П., Масели Э., Джонсон Дж. И др. (2012). Геномные основы адаптивной эволюции трехиглой колюшки. Природа 484 55–61. 10.1038 / nature10944 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Камберов Ю.G., Wang S., Tan J., Gerbault P., Wark A., Tan L. и др. (2013). Моделирование недавней эволюции человека на мышах путем экспрессии выбранного варианта EDAR. Ячейка 152 691–702. 10.1016 / j.cell.2013.01.016 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Kammenga JE, Doroszuk A., Riksen JAG, Hazendonk E., Spiridon L., Petrescu A.-J. , и другие. (2007). Caenorhabditis elegans дикого типа не подчиняется правилу температуры и размера из-за однонуклеотидного полиморфизма в tra-3. PLoS Genet 3: e34 10.1371 / journal.pgen.0030034 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Keller E.F. (2010). Мираж пространства между природой и заботой .
Дарем, Северная Каролина: издательство Duke University Press;
10.1215 / 97808223
[CrossRef] [Google Scholar]
- Киршнер М., Герхарт Дж. (1998). Эволюционируемость. Proc. Natl. Акад. Sci. США 95 8420–8427. 10.1073 / pnas.95.15.8420 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Кислухин Г., Кинг Э. Г., Уолтерс К. Н., Макдональд С. Дж., Лонг А. Д. (2013). Генетическая архитектура токсичности метотрексата сходна у Drosophila melanogaster и человека. 3 1301–1310. 10.1534 / g3.113.006619 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ландри К. Р., Рифкин С. А. (2012). Карты генотипа-фенотипа системной биологии и количественной генетики: отдельные и дополняющие друг друга. Adv. Exp. Med. Биол. 751 371–398. 10.1007 / 978-1-4614-3567-9_17 [PubMed] [CrossRef] [Google Scholar]
- Langlade N.Б., Фэн Х., Дрансфилд Т., Копси Л., Ханна А. И., Тебо К. и др. (2005). Эволюция через генетически контролируемое пространство аллометрии. Proc. Natl. Акад. Sci. США 102 10221–10226. 10.1073 / pnas.0504210102 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Lenser T., Theißen G. (2013). Молекулярные механизмы, участвующие в одомашнивании конвергентных культур. Trends Plant Sci. 18 704–714. 10.1016 / j.tplants.2013.08.007 [PubMed] [CrossRef] [Google Scholar]
- Lettre G.(2011). Последние успехи в изучении генетики роста. Hum. Genet. 129 465–472. 10.1007 / s00439-011-0969-x [PubMed] [CrossRef] [Google Scholar]
- Левонтин Р. (1974a). Генетическая основа эволюционных изменений (Колумбийская биологическая серия) . Колумбия: Издательство Колумбийского университета. [Google Scholar]
- Левонтин Р. К. (1974b). Дисперсионный анализ и анализ причин. г. J. Hum. Genet. 26 400–411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Левонтин Р.(1978). Приспособление. Sci. Являюсь. 293 212–228. 10.1038 / Scientificamerican0978-212 [PubMed] [CrossRef] [Google Scholar]
- Линнен К. Р., По Й.-П., Петерсон Б. К., Барретт Р. Д. Х., Ларсон Дж. Г., Дженсен Дж. Д. и др. (2013). Адаптивная эволюция множества признаков посредством множественных мутаций одного гена. Наука 339 1312–1316. 10.1126 / science.1233213 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Лю Ф., Вэнь Б., Кайзер М. (2013). Цветные полиморфизмы ДНК человека. Семин. Cell Dev. Биол. 24 562–575. 10.1016 / j.semcdb.2013.03.013 [PubMed] [CrossRef] [Google Scholar]
- Луонг Х. Т., Чаплин Дж., Макрей А. Ф., Медланд С. Э., Виллемсен Г., Найхолт Д. Р. и др. (2011). Вариация BMPR1B, TGFRB1 и BMPR2 и контроль дизиготного двойникования. Twin Res. Гм. Genet. Выключенный. J. Int. Soc. Twin Stud. 14 408–416. 10.1375 / twin.14.5.408 [PubMed] [CrossRef] [Google Scholar]
- Линч М., Уолш Б. (1998). Генетика и анализ количественных признаков .Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
- MacCallum D. M., Coste A., Ischer F., Jacobsen M. D., Odds F. C., Sanglard D. (2010). Генетическое вскрытие механизмов устойчивости к азолам у Candida albicans и их проверка на мышиной модели диссеминированной инфекции. Антимикробный. Агенты Chemother. 54 1476–1483. 10.1128 / AAC.01645-1649 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Манске М., Миотто О., Кампино С., Оберн С., Альмагро-Гарсия Дж., Маслен Г. и др. (2012). Анализ разнообразия Plasmodium falciparum естественных инфекций методом глубокого секвенирования. Природа 487 375–379. 10.1038 / nature11174 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Мартин А., Оргогозо В. (2013a). Данные из: Локусы повторной эволюции: Каталог генетических горячих точек фенотипической изменчивости . Доступно на: http://datadryad.org/resource/ 10.5061 / dryad.v66p0 [PubMed] [CrossRef] [Google Scholar]
- Мартин А., Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических горячих точек фенотипической изменчивости. Evol. Int. J. Orgn. Evol. 67 1235–1250. 10.1111 / evo.12081 [PubMed] [CrossRef] [Google Scholar]
- Mayr E. (1963). Виды животных и эволюция . Гарвард: издательство Гарвардского университета; 10.4159 / harvard.9780674865327 [CrossRef] [Google Scholar]
- МакГэри К. Л., Парк Т. Дж., Вудс Дж. О., Ча Х. Дж., Уоллингфорд Дж. Б., Маркотт Э. М. (2010). Систематическое открытие неочевидных моделей болезней человека через ортологичные фенотипы. Proc. Natl. Акад. Sci. США 107 6544–6549. 10.1073 / pnas.00107 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Mery F., Belay A. T., So A. K.-C., Sokolowski M. B., Kawecki T. J. (2007). Естественный полиморфизм, влияющий на обучение и память у Drosophila . Proc. Natl. Акад. Sci. США 104 13051–13055. 10.1073 / pnas.07024 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Moss L. (2003). Один, два (тоже?), Много генов? В.Rev. Biol. 78 57–67. 10.1086 / 367581 [CrossRef] [Google Scholar]
- Народ С. А., Фоулкс В. Д. (2004). BRCA1 и BRCA2: 1994 и позже. Nat. Преподобный Рак 4 665–676. 10.1038 / nrc1431 [PubMed] [CrossRef] [Google Scholar]
- Натараджан К., Иногучи Н., Вебер Р. Э., Фаго А., Морияма Х., Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340 1324–1327. 10.1126 / science.1236862 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Noble D.(2006). Музыка жизни: биология вне генома . Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- Осборн К. А., Робишон А., Берджесс Э., Батланд С., Шоу Р. А., Култхард А. и др. (1997). Полиморфизм естественного поведения, обусловленный цГМФ-зависимой протеинкиназой Drosophila . Наука 277 834–836. 10.1126 / science.277.5327.834 [PubMed] [CrossRef] [Google Scholar]
- Ояма С. (2000). Онтогенез информации: системы развития и эволюция 2-е изд.Кембридж, Нью-Йорк: издательство Duke University Press; 10.1215 / 9780822380665 [CrossRef] [Google Scholar]
- Пааби А. Б., Рокман М. В. (2013). Многоликая плейотропия. Trends Genet. 29 66–73. 10.1016 / j.tig.2012.10.010 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Палмер А. Р. (2004). Нарушение симметрии и эволюция развития. Наука 306 828–833. 10.1126 / science.1103707 [PubMed] [CrossRef] [Google Scholar]
- Патерсон А. Х., Лин Ю.Р., Ли З., Шерц К. Ф., Добли Дж. Ф., Пинсон С. Р. и др. (1995). Конвергентное одомашнивание зерновых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269 1714–1718 гг. 10.1126 / science.269.5231.1714 [PubMed] [CrossRef] [Google Scholar]
- Perutz M. F. (1983). Адаптация видов в белковой молекуле. Мол. Биол. Evol. 1 1–28. [PubMed] [Google Scholar]
- Филлипс П. К. (2008). Эпистаз — важнейшая роль взаимодействия генов в структуре и эволюции генетических систем. Nat. Преподобный Жене. 9 855–867. 10.1038 / nrg2452 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Pokorná M., Kratochvíl L. (2009). Филогения механизмов определения пола у плоских рептилий: являются ли половые хромосомы эволюционной ловушкой? Zool. J. Linn. Soc. 156 168–183. 10.1111 / j.1096-3642.2008.00481.x [CrossRef] [Google Scholar]
- Робинсон П. Н., Уэббер К. (2014). Онтологии фенотипов и межвидовой анализ для трансляционных исследований. PLoS Genet 10: e1004268 10.1371 / journal.pgen.1004268 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Rockman M. V. (2012). Программа QTN и аллели, имеющие значение для эволюции: все это золото не блестит. Evol. Int. J. Orgn. Evol. 66 1–17. 10.1111 / j.1558-5646.2011.01486.x [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Роджерс В. А., Саломон Дж. Р., Тейси Д. Дж., Камино Э. М., Дэвис К. А., Ребеиз М. и др. (2013). В основе разнообразия пигментации плодовой мушки лежит рекуррентная модификация консервативного цис-регуляторного элемента. PLoS Genet 9: e1003740 10.1371 / journal.pgen.1003740 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ромеро-Эррера А. Э., Леманн Х., Джойси К. А., Пятница А. Э. (1978). Об эволюции миоглобина. Philos. Пер. R. Soc. Лондон. B. Biol. Sci. 283 61–163. 10.1098 / rstb.1978.0018 [PubMed] [CrossRef] [Google Scholar]
- Салазар-Сьюдад И., Марин-Риера М. (2013). Адаптивная динамика в картах генотип-фенотип на основе развития. Природа 497 361–364.10.1038 / nature12142 [PubMed] [CrossRef] [Google Scholar]
- Саркар С. (1999). От нормы реакции к адаптивной норме: норма реакции, 1909–1960 гг. Biol. Филос. 14 235–252. 10.1023 / A: 10066648 [CrossRef] [Google Scholar]
- Швандер Т., Леймар О. (2011). Гены как лидеры и последователи эволюции. Trends Ecol. Evol. 26 143–151. 10.1016 / j.tree.2010.12.010 [PubMed] [CrossRef] [Google Scholar]
- Schwartz S. (2000). «Дифференциальная концепция гена: прошлое и настоящее», в Концепция гена в развитии и эволюции редакторы Бертон П.Дж., Фальк Р., Райнбергер Х. Дж. (Кембридж, Нью-Йорк: издательство Кембриджского университета;) 26–39. 10.1017 / CBO9780511527296.004 [CrossRef] [Google Scholar]
- Ширанги Т. Р., Дюфур Х. Д., Уильямс Т. М., Кэрролл С. Б. (2009). Быстрая эволюция экспрессии фермента, продуцирующего половые феромоны, у Drosophila . PLoS Biol. 7: e1000168 10.1371 / journal.pbio.1000168 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Симондон Г. (1968). L’individuation à la Lumière des Notions de Forme et d’Information 2005-е изд.Гренобль: Миллон. [Google Scholar]
- Собер Э. (1988). Распределение причинно-следственной ответственности. J. Philos. 85 303–318. 10.2307 / 2026721 [CrossRef] [Google Scholar]
- Штайнер К. К., Вебер Дж. Н., Хекстра Х. Э. (2007). Адаптивная изменчивость пляжных мышей вызвана двумя взаимодействующими генами пигментации. PLoS Biol. 5: e219 10.1371 / journal.pbio.0050219 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Стерн Д. Л. (2000). Эволюционная биология развития и проблема вариации. Evol. Int. J. Orgn. Evol. 54 1079–1091. 10.1111 / j.0014-3820.2000.tb00544.x [PubMed] [CrossRef] [Google Scholar]
- Стерн Д. Л. (2013). Генетические причины конвергентной эволюции. Nat. Преподобный Жене. 14 751–764. 10.1038 / nrg3483 [PubMed] [CrossRef] [Google Scholar]
- Стерн Д., Оргогозо В. (2008). Локусы эволюции: насколько предсказуема генетическая эволюция? Evol. Int. J. Orgn. Evol. 62 2155–2177. 10.1111 / j.1558-5646.2008.00450.x [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Stewart C.Б., Шиллинг Дж. У., Уилсон А. С. (1987). Адаптивная эволюция лизоцимов желудка ферментеров передней кишки. Природа 330 401–404. 10.1038 / 330401a0 [PubMed] [CrossRef] [Google Scholar]
- Stotz K. (2012). Убийство на экспрессе развития: кто убил природу / воспитание? Биол. Philos. 27 919–929. 10.1007 / s10539-012-9343-1 [CrossRef] [Google Scholar]
- Стертевант А. Х. (1932). Использование мозаики в изучении влияния генов на развитие. Proc.Шестой Int. Congr. Genet. Итака Н. Я. 1 304–307. [Google Scholar]
- Supple M., Papa R., Counterman B., McMillan W. O. (2014). Геномика адаптивного излучения: понимание континуума видообразования геликониусов. Adv. Exp. Med. Биол . 781 249–271. 10.1007 / 978-94-007-7347-9_13 [PubMed] [CrossRef] [Google Scholar]
- Tautz D., Schmid K. J. (1998). От генов к людям: гены развития и формирование фенотипа. Philos. Пер. Р.Soc. Лондон. B. Biol. Sci. 353 231–240. 10.1098 / rstb.1998.0205 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Thomas D. (2010). Исследования ассоциации генов и окружающей среды: новые подходы. Nat. Преподобный Жене. 11 259–272. 10.1038 / nrg2764 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Венкатеш Б., Лу С. К., Дандона Н., См. С. Л., Бреннер С., Сунг Т. В. (2005). Генетические основы устойчивости к тетродотоксинам у иглобрюхов. Curr. Биол. 15 2069–2072. 10.1016 / j.cub.2005.10.068 [PubMed] [CrossRef] [Google Scholar]
- Висшер П. М., Браун М. А., Маккарти М. И., Ян Дж. (2012). Пять лет открытия GWAS. г. J. Hum. Genet. 90 7–24. 10.1016 / j.ajhg.2011.11.029 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Waddington C.H. (1957). Стратегия генов 2014 Эдн Перепечатка Лондон: Рутледж. [Google Scholar]
- Вагнер Г. П. (1996). Гомологи, естественные виды и эволюция модульности. г. Zool. 36 36–43. 10.1093 / icb / 36.1.36 [CrossRef] [Google Scholar]
- Вагнер Г. П. (2000). Концепция персонажа в эволюционной биологии . Сан-Диего, Калифорния: Academic Press. [Google Scholar]
- Вагнер Г. П. (2014). Гомология, гены и эволюционные инновации . Принстон, Нью-Джерси: Издательство Принстонского университета; 10.1515 / 9781400851461 [CrossRef] [Google Scholar]
- Wang J., Wurm Y., Nipitwattanaphon M., Riba-Grognuz O., Huang Y.-C., Shoemaker D., и другие. (2013). Y-подобная социальная хромосома вызывает альтернативную организацию колоний у огненных муравьев. Природа 493 664–668. 10.1038 / nature11832 [PubMed] [CrossRef] [Google Scholar]
- Уотерс К. К. (2007). Причины, которые имеют значение. J. Philos. 104 551–579. [Google Scholar]
- Велтер Д., МакАртур Дж., Моралес Дж., Бёрдетт Т., Холл П., Джанкинс Х. и др. (2014). Каталог NHGRI GWAS, кураторский ресурс ассоциаций SNP-признаков. Nucleic Acids Res. 42 D1001 – D1006. 10.1093 / nar / gkt1229 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Вест-Эберхард М. Дж. (2003). Пластичность и эволюция развития . Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- Вест-Эберхард М. Дж. (2005). Пластичность развития и происхождение видовых различий. Proc. Natl. Акад. Sci. США 102 (Дополнение 1) 6543–6549. 10.1073 / pnas.0501844102 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Wilkins A.(2014). «Набор генетических инструментов»: история жизни важной метафоры », в Достижения в эволюционной биологии развития изд. Тодд Стрелман Дж. (Хобокен, Нью-Джерси: John Wiley & Sons;) 1–14. [Google Scholar]
- Уилкинс А. С. (2002). Эволюция путей развития . Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
- Xu X., Dong G.-X., Hu X.-S., Miao L., Zhang X.-L., Zhang D.-L., et al. (2013). Генетическая основа белых тигров. Curr.Биол. 23 1031–1035. 10.1016 / j.cub.2013.04.054 [PubMed] [CrossRef] [Google Scholar]
- Чжан Дж., Вагнер Г. П. (2013). Об определении и измерении плейотропии. Trends Genet. 29 383–384. 10.1016 / j.tig.2013.05.002 [PubMed] [CrossRef] [Google Scholar]
Реакция пластика на новую среду смещена в сторону размеров фенотипа с высокой аддитивной генетической изменчивостью
Значимость
Теория предполагает, что популяции могут развиваться генетически. модификация реакций, вызванных окружающей средой.Мы демонстрируем, что такие пластичные реакции на новую среду имеют тенденцию происходить по признакам, имеющим высокую генетическую изменчивость. Это предполагает, что отбор за или против фенотипов, индуцированных окружающей средой, обычно будет эффективным. То, что организмы одинаково реагируют на новизну окружающей среды и генетические вариации, можно объяснить тем, как устроено развитие. Это затрудняет определение того, формируют ли пластичность или генетические ограничения эволюционные реакции на изменение окружающей среды.
Abstract
Фенотипы, вызванные окружающей средой, были предложены для инициирования и смещения адаптивных эволюционных изменений в определенных направлениях. Вероятность того, что это произойдет, частично зависит от того, насколько хорошо пластические реакции согласуются с аддитивной генетической дисперсией и ковариацией признаков. Используя метаанализ, мы демонстрируем, что пластическая реакция на новую среду имеет тенденцию происходить по фенотипическим измерениям, которые несут в себе существенное количество аддитивных генетических вариаций.Это предполагает, что отбор за или против фенотипов, индуцированных окружающей средой, обычно будет эффективным. Одна интерпретация соответствия между направлением пластичности и главной осью аддитивной генетической изменчивости состоит в том, что системы развития имеют тенденцию реагировать на новизну окружающей среды так же, как и на генетические мутации. Это затрудняет определение того, определяется ли направление эволюции пластичностью или генетическим «ограничением». Таким образом, наши результаты подчеркивают необходимость новых теоретических и эмпирических подходов к рассмотрению роли пластичности в эволюции.
Популяции, которые сталкиваются с новой средой, часто должны адаптироваться, чтобы выжить. Скорость и направление адаптивных изменений зависят от фенотипических и генетических вариаций, которые подвергаются естественному отбору. Гипотеза «прежде всего пластичность» предполагает, что обусловленные окружающей средой фенотипические вариации инициируют и направляют адаптивную генетическую дивергенцию по определенным траекториям (1, 2). Это было продемонстрировано экспериментально (3, 4), и наблюдение, что пластические реакции иногда совпадают с популяционной дивергенцией по морфологии, физиологии и поведению, делает это правдоподобным объяснением адаптивной дивергенции в дикой природе (5-10). .Однако, склонна ли эволюционная дивергенция в целом следовать направлению фенотипов, индуцированных окружающей средой, оспаривается и плохо понимается (рассмотрено в ссылках 2 и 11–14).
Предполагается, что адаптация после воздействия новой или стрессовой среды включает в себя уточнение, преувеличение или устранение пластичных реакций предков посредством количественных генетических изменений. Это возможно только тогда, когда есть наследственные вариации по размерам фенотипа, которые являются пластичными.В количественной генетике эта вариация описывается в терминах аддитивной генетической дисперсии и ковариации между признаками, обычно называемой матрицей G . Прогнозируемое изменение фенотипа при направленном отборе на постоянную генетическую изменчивость считается количественной генетической оценкой эволюционируемости (15). Количественная генетическая эволюционирующая способность наиболее высока в направлении фенотипического пространства с наиболее аддитивной генетической изменчивостью. При прочих равных, количественная генетическая эволюционируемость фенотипов, индуцированных окружающей средой, поэтому будет высокой, когда направление фенотипической пластичности хорошо выровнено с осью максимальной аддитивной генетической изменчивости в новых средах.Мы называем это выравнивание краткосрочным эволюционным потенциалом (ссылка 1, стр. 142) вызванных окружающей средой фенотипов или пластических реакций.
Есть основания полагать, что направление пластичности и главная ось генетической изменчивости могут совпадать. Как генетические, так и средовые возмущения вызывают свои фенотипические эффекты в процессе развития, а фенотипы, вызванные мутацией, часто также могут быть вызваны факторами окружающей среды, и наоборот (1). Матрица G частично является результатом распределения мутационных эффектов (в дополнение к, e.g., выборочная потеря вариативности) и, следовательно, будет иметь тенденцию отражать, как черты регулируются в процессе развития (16). В самом деле, модели демонстрируют, что корреляционный отбор может производить взаимодействия в процессе развития, которые направляют фенотипические эффекты мутации по размерам комбинаций признаков, которые были предпочтительны в прошлом (17-21). Поскольку фенотипическая аккомодация включает скоординированную настройку признаков, которые должны функционировать вместе (1), она также зависит от взаимодействий в процессе развития, которые, вероятно, являются продуктами корреляционного отбора.Таким образом, отбор по фенотипам, индуцированным окружающей средой, может изменить регуляцию фенотипов в развитии таким образом, что он увеличивает аддитивную генетическую изменчивость по размерам фенотипа, которые являются пластичными (22). Если это так, это означает, что отбор в направлении или противодействие пластической реакции на новую среду обычно будет эффективным, что может предрасполагать популяцию к краткосрочному развитию в соответствии с измерениями фенотипа, которые являются пластичными.
Здесь мы используем метаанализ, чтобы проверить, смещены ли пластические реакции в сторону размеров черт, которые несут высокий уровень аддитивной генетической изменчивости.Пластические реакции можно описать с помощью «вектора пластичности», который количественно определяет изменение средних значений фенотипических признаков между двумя средами, выраженное как расстояние между средними значениями многомерного фенотипа. Мы решили сосредоточиться на условиях окружающей среды, к которым популяция (пока) не адаптирована, поскольку это наиболее спорный сценарий эволюции, обусловленной пластичностью. Потенциал адаптивного расхождения по измерениям признаков, которые являются экологически чувствительными, будет высоким, когда (–) генетическая вариация, лежащая в основе фенотипической дисперсии и ковариации (матрица P ), увеличивается в новой среде и ( ii ) пластическая реакция («Вектор пластичности») падает вдоль главной оси G , в то время как ориентация G остается неизменной в новой среде, или ( iii ) пластический отклик не падает вдоль главной оси G , но ориентация G поворачивается в новой среде, так что она совпадает с направлением пластичности (рис.1). Наш мета-анализ проверяет, насколько распространены эти сценарии.
Рис. 1.Альтернативные сценарии, описывающие различия в краткосрочном эволюционном потенциале экологических фенотипов. Эллипсы отображают многомерные фенотипические ( P , светлый цвет) и генотипические ( G , темные цвета) вариации для двух признаков (Признак 1 и Признак 2). Стрелки, соединяющие центроиды необычной (зеленый) и новой (оранжевый) сред, являются векторами пластичности, которые описывают изменение многомерного среднего фенотипа ( P ) между двумя средами.( A ) Можно сказать, что пластическая реакция на новую среду имеет высокий эволюционный потенциал, если существует обширная генетическая изменчивость, лежащая в основе P , и вектор пластической реакции хорошо согласован с главной осью генетической изменчивости в новой среде ( т.е. темно-оранжевый эллипс). ( B ) Можно сказать, что пластиковый фенотипический ответ имеет низкий эволюционный потенциал, если существует ограниченная генетическая изменчивость, лежащая в основе P , и вектор пластического ответа не совпадает с главной осью генетической изменчивости в новой среде.
Результаты
Мы провели систематический поиск литературы (более подробно см. Материалы и методы ) и выявили 21 исследование по 19 видам, в которых можно было сравнить матрицы G и P (т. Е. 32 экологических сравнения G и P с n = всего 64 эффекта). Мы также идентифицировали 32 исследования ( n = 98 эффектов) по 30 видам, которые можно было бы использовать для получения оценок количественной генетической эволюционируемости в направлении пластичности.Это большее количество исследований могло быть включено, потому что нам нужен был только вектор средних признаков и матрицы G для получения размеров эффекта (см. Материалы и методы для более подробной информации). Данные были таксономически смещены в сторону насекомых с общим количеством эффектов 56, тогда как рыбы и растения имели 18 и 16 размеров эффекта соответственно. Были также оценены матрицы G для одного вида улиток и одного вида лягушек (с шестью и двумя эффектами соответственно). Размер матрицы варьировал от 2 до 11 признаков (среднее значение = 4.16, n = 98), и новые условия были введены в условиях, которые были либо «стрессовыми» (приводящими к снижению выживаемости), либо «нестрессовыми», и с использованием различных схем разведения. Учитывая, что эти факторы могут повлиять на G (23⇓ – 25), мы учли их в нашем анализе (см. Ниже и Материалы и методы для более подробной информации).
Изменения аддитивной генетической дисперсии в ответ на новые среды.
Сначала мы исследовали, приводят ли новые условия окружающей среды к увеличению аддитивной генетической изменчивости (т.е., «загадочная генетическая изменчивость»; ссылки 23 и 24), создавая размеры эффектов, которые контрастировали структуру G между непривычной и новой средами. Не новаторская среда — это та среда, к которой популяции считались хорошо адаптированными или в которой популяции выращивались не менее пяти поколений или более, тогда как новой средой считались те, с которыми манипулировали вне нормальных условий выращивания. Мы количественно оценили изменения в аддитивной генетической изменчивости, рассчитав количество величин эффекта, описывающих изменения в G в разных средах (см. Материалы и методы для более подробной информации).Здесь мы сосредотачиваемся на тех, которые описывают изменение общей стандартизированной генетической изменчивости (SDV) и изменение доли аддитивной генетической изменчивости вдоль главной оси G (т.е. выпущен в ответ на новые условия. В некоторых исследованиях новая среда считалась стрессовой и / или приводила к снижению выживаемости (см. Материалы и методы ). Поэтому, используя модель метарегрессии, мы проверили, зависели ли наблюдаемые изменения в G от того, была ли новая среда «стрессовой» или «нестрессовой».Мы также проверили, повлияли ли тип разведения («полусиб» или «полусиб») и количество признаков на величину изменений.
В разных исследованиях было мало доказательств высвобождения загадочной генетической изменчивости (общее метааналитическое среднее для моделей только с перехватом: SDV: = -0,12, 95% ДИ = -0,47 до 0,23; PVmax = -0,06, 95% CI = от -0,68 до 0,55). Однако наблюдалась большая разница между исследованиями в величине эффекта (доля дисперсии между исследованиями [Ist2]: SDV: = 0.84; PVmax = 0,73). При контроле количества признаков, дизайна разведения и того, были ли манипуляции с окружающей средой стрессовыми, были доказательства того, что дисперсия может увеличиваться в новых условиях. В частности, дисперсия признаков значительно увеличилась в новых условиях для исследований с использованием планов разведения полных сибсов [SDV: общая средняя оценка (логит-шкала) = 0,62, 95% доверительный интервал = 0,086–1,159; Рис. 2 A и см. SI Приложение , Таблица S2], где ∼61% общей аддитивной генетической изменчивости приходится на gmax [PVmax: общая средняя оценка (логит-шкала) = 0.465, 95% ДИ = от -0,5 до 1,43; Рис. 2 B и см. SI Приложение , Таблица S2]. Тем не менее, матрицы G , оцененные с использованием планов разведения полусибсов, на самом деле показали более низкую общую аддитивную генетическую дисперсию в новых средах и более низкую долю аддитивной генетической дисперсии по gmax по сравнению с исследованиями с использованием планов разведения полных сибсов (на ~ 35%; рис. 2 A и B ). Была ли новая среда стрессовой или нет, это не повлияло на изменение аддитивной генетической изменчивости (рис.2 A и B ).
Рис. 2.Средние величины эффекта, сравнивающие изменение генетической изменчивости в разных средах для исследований, в которых новые манипуляции с окружающей средой были стрессовыми (стрессовые / нестрессовые) и с различными схемами разведения (полусиб против полных сестер). ( A ) Общий SDV между матрицами G в разных средах. ( B ) Изменение доли вариации по gmax (PV max ) между матрицами G в разных средах.Обратите внимание, что во всех случаях величина эффекта является средним значением для исследования среднего числа признаков и не зависит от других факторов (т. Е. Маргинальных средних) в модели (см. Материалы и методы ).
Оценка краткосрочного эволюционного потенциала экологически индуцированных фенотипов.
Потенциал обусловленных окружающей средой фенотипов смещать эволюционную дивергенцию зависит от степени, в которой пластические реакции попадают в размерности фенотипа, который несет наследственную изменчивость (рис.1 А ). Мы получили новые меры величины эффекта, вдохновленные Хансеном и Хоулом (15), чтобы проверить это предсказание. Сначала мы количественно оценили, насколько аддитивная генетическая изменчивость упала в направлении пластической реакции (определяемой как πe), а затем вывели меру количественной генетической эволюционируемости ( sensu , ссылка 15), определяемую как выравнивание между вектором пластической реакции и основным оси аддитивной генетической изменчивости (gmax) как в новой, так и в новой среде (обозначается как θe; подробности см. в материалах и методах ).Мы также исследовали взаимосвязь между G и P внутри и между средами и представили эти результаты в приложении SI , таблицах S2 и S3. Мы явно проверили, произошли ли существенные изменения в том, как вектор пластической реакции согласовывался с аддитивной генетической изменчивостью, когда популяция переходит из не новаторской среды в новую, с использованием модели метарегрессии, противопоставляющей тип среды (не новаторский и новый), после контроля количества признаков и разведения дизайн.
В среднем, наблюдалась существенная аддитивная генетическая дисперсия в признаках вдоль вектора пластичности как в не нововведении, так и в новой среде (средняя дисперсия 69,14% в не новелле и 71,06% в романе; Рис.3 A и SI Приложение , таблица S3). Пластические ответы падали между 32 и 42 ° от главной оси аддитивной генетической изменчивости (т.е. gmax x ; Рис. 3 B и см. SI Приложение , Таблица S3). Это выравнивание между вектором пластичности и gmax было значительно больше, чем ожидалось при случайном выравнивании ( SI, приложение , таблица S1).
Рис. 3.Средние величины эффекта при сравнении пластических реакций на G в не новаторских и новых средах (Nonnovel vs.Roman) и с использованием различных схем разведения (Half-sib против Full-sib). ( A ) Доля генетической изменчивости по вектору пластической реакции (πe). ( B ) Выравнивание между вектором пластического ответа и основными осями генетической изменчивости gmax (θe). Отрицательные значения указывают на то, что вектор пластической реакции и gmax полностью выровнены (т. Е. ∼0 °), тогда как положительные значения указывают на то, что вектор пластической реакции и gmax не выровнены (т.е.э., ∼90 °). Значения 0 указывают на то, что вектор пластической реакции и gmax находятся под углом 45 ° друг к другу. Обратите внимание, что во всех случаях величина эффекта является средним значением для исследования среднего количества признаков и не зависит от других факторов (т. Е. Маргинальных средних) в модели ( Материалы и методы, ).
Однако угол между вектором пластического отклика и gmax (т. Е. Θe) для не новаторских и новых сред существенно варьировался в разных исследованиях (неоднородность между исследованиями; Ist2 = 0.80; Рис.4). Эти результаты показывают, что результаты зависят от типов измеряемых характеристик, типа воздействия окружающей среды и / или прошлой выборочной истории; Однако выявить эти источники вариации невозможно с существующими данными. В некоторых исследованиях вращение матрицы G заставляло вектор пластичности выравниваться лучше в новой среде, чем в не новой среде, тогда как в других исследованиях происходило противоположное (рис. 5).
Рис. 4.( A ) Общее количество генетических вариаций вдоль вектора пластической реакции как пропорция максимального количества вариаций в любом направлении, πe.Меньшие значения на осях (т. Е. Отрицательные числа) соответствуют ситуациям, когда генетическая изменчивость по вектору пластичности низкая (метка «Низкая»), тогда как большие положительные значения соответствуют ситуациям, когда генетическая изменчивость по вектору пластичности является высокой (метка «Высокая» ) для не новеллы (зеленый текст) и романа (оранжевый текст). ( B ) Угол между вектором пластической реакции и gmax (θe) в новых и не новаторских условиях во всех исследованиях. Метки соответствуют значениям на осях, где вектор пластичности не совпадает с главной осью генетической изменчивости gmax (плохо выровнен) или где gmax совпадает с вектором пластичности (хорошо выровнен) как в романе (зеленый текст), так и в романе. (оранжевый текст) среды.Оси нанесены на преобразованную шкалу, чтобы облегчить сравнение с результатами анализа. Большие отрицательные значения (например, -8) указывают на то, что вектор пластической реакции и gmax полностью выровнены (т. Е. ∼0 °), тогда как большие положительные значения (например, 8) указывают, что вектор пластической реакции и gmax не выровнены (т. Е. ∼90 °). Значения 0 указывают на то, что вектор пластической реакции и gmax находятся под углом 45 ° друг к другу. Показаны средние размеры эффекта и 95% интервалы выборки.
Рис. 5.Примеры исследований (с двумя матрицами признаков) с высоким ( A и B ) или низким ( C и D ) выравниванием между направлением пластических ответов и постоянной генетической изменчивостью.Стрелки — это пластиковые векторы отклика; большие светлые эллипсы — это матрицы P , а меньшие темные эллипсы — это матрицы G . Цвета соответствуют неровной (зеленый) и новой (оранжевый) средам. ( A ) Kasule (44) использовал клопа для окрашивания хлопка ( Dysdercus fasciatus ) для оценки количественной генетической изменчивости плодовитости и размера взрослых особей в стрессовых условиях с использованием схемы разведения полусибсов. ( B ) Blanckenhorn и Heyland (45) использовали план разведения полных сибсов для оценки количественной генетической изменчивости размера и времени развития задних конечностей у желтой навозной мухи ( Scathophaga stercoraria ) в стрессовых условиях.( C ) Виа и Коннер (46) изучали мучного жука ( Tribolium castaneum ), используя схему разведения полусибсов, чтобы оценить количественные генетические вариации веса куколки и времени развития в нестрессовых условиях. ( D ) Каузе и Морин (47) изучили пилильщика ( Priophorus pallipes ) и оценили количественные генетические параметры для времени развития и размера тела, используя метод разведения полных сибсов в стрессовых условиях окружающей среды.
Обсуждение
Вест-Эберхард предположил, что «гены являются последователями, а не лидерами в адаптивной эволюции» (1).Эволюционное преувеличение или уменьшение фенотипов, вызванных окружающей средой, требует наследственных вариаций по размерам фенотипа, которые являются пластичными. Наши результаты демонстрируют, что соответствие между пластическими реакциями на новую среду и основной осью генетической изменчивости больше, чем ожидалось случайно (Рис. 4 и SI Приложение , Таблица S1). Это может отражать системы развития, аналогично реагирующие на факторы окружающей среды и генетические факторы. Такие отклонения в развитии должны способствовать адаптивной дивергенции в ответ на отбор по фенотипам, вызванным окружающей средой, благодаря чему пластичность, по-видимому, играет ведущую роль в адаптивной эволюции (1).
Не совсем понятно, почему генетические и экологические нарушения должны иметь сходные фенотипические эффекты. Однако теоретические модели продемонстрировали, что развитые регуляторные системы имеют тенденцию производить ошибки развития, которые отражают корреляционный отбор в прошлом (рассмотрено в ссылках 20 и 26). Например, многовариантный отбор может предрасполагать системы развития к пластической реакции в направлении ковариации признаков (22). Подобные предубеждения в фенотипах могут возникать в ответ на мутации (например,г., исх. 21). Матрица мутационной дисперсии и ковариации (матрица M ; ссылки 27 и 28) является более подходящей оценкой смещения развития, чем G , поскольку последняя также отражает эффекты отбора и дрейфа. Тем не менее, G часто предполагается, что представляет, как фенотипы изменяются в ответ на генетические вариации (например, ссылки 29 и 30). Другими словами, поскольку и направление пластичности, и матрица G фиксируют, как регулируется развитие признаков, пластические реакции и основная ось аддитивной генетической изменчивости обычно могут быть совмещены.Наши результаты показывают, что это действительно может иметь место для многих черт, даже для пластиковой реакции на среду, к которой организмы не адаптированы.
Воздействие новой или стрессовой среды также может быть связано с общим увеличением аддитивной генетической изменчивости (23). Высвобождение «загадочной генетической изменчивости» иногда считается центральным в моделях эволюции, ориентированных на пластичность (2, 31). В то время как аддитивная генетическая изменчивость обычно увеличивалась в новых средах, исследования с использованием схем разведения полукровок фактически подтвердили уменьшение аддитивной генетической изменчивости.Это различие между схемами разведения предполагает, что материнские или отцовские эффекты вносят значительный вклад в фенотипические вариации, которые проявляются в новых средах, и что вариативность, вносимая родительскими эффектами, может привести к лучшему согласованию с пластической реакцией. Этот результат обеспечивает более детальное понимание предыдущей работы; без эффективного разделения на окружающую среду и генетические эффекты исследования могут переоценить, насколько генетическая изменчивость является «загадочной» (32, 33).
Хотя эти результаты предполагают существенные возможности для дивергенции популяций в направлении пластичности, влияние пластичности на эволюционные траектории зависит от силы и формы отбора, действующего на распределение фенотипов в новых средах — информация, которая часто отсутствует. Например, исследования в нашем метаанализе редко оценивали отбор по фенотипам в новой среде (это также означает, что мы не могли учесть какой-либо отбор, который мог произойти из-за смертности).Это неудивительно, учитывая, что оценка отбора требует дополнительных или более сложных экспериментов. На сегодняшний день только одно исследование явно рассматривало соответствие между gmax, пластичностью и отбором (34). Lind et al. (34) показали, что у водяных блох адаптивная пластичность в ответ на сигналы рыб-хищников хорошо согласуется с gmax и реакцией на выбор выживаемости; однако то же самое было не так в ответ на реплики хищников мошек. Неясно, связано ли это различие с силой прошлой селекции, но клоны водяных блох считались хорошо адаптированными к обоим режимам хищников (вот почему исследование не было включено в наш метаанализ).
То, что эволюционные реакции нелегко предсказать на основе отбора или изменчивости, также очевидно из компиляции 16 долгосрочных исследуемых популяций, которые показали, что по мере изменения окружающей среды взаимосвязь между наследственной изменчивостью отдельных признаков (т. Е. h 2 ) и слабый отбор (35). Действительно, некоторые исследования в настоящем анализе показали, что направление пластичности и постоянная генетическая изменчивость плохо согласованы. На основе теоретических моделей, описанных выше, может показаться разумным заключить, что такие случаи плохого выравнивания просто отражают то, что люди не могут успешно приспособиться к новым условиям.Однако эта интерпретация осложняется не в последнюю очередь тем, что G также отражает избирательное удаление генетической изменчивости (36). Это означает, что большая часть аддитивной генетической изменчивости, обнаруженная исследованиями в этом метаанализе, могла быть выборочно нейтральной в среде предков. Если это так, то соответствие между аддитивной генетической дисперсией и направлением пластичности можно интерпретировать так, как будто пластичность смещена в сторону размеров фенотипа, которые были слабо, а не сильно отобраны в прошлом.Эта неоднозначность в интерпретации соответствия между пластичными ответами и G требует теоретических и эмпирических исследований, которые могут продемонстрировать, как эволюция развития формирует пластичность и постоянную генетическую изменчивость, последнее — через фенотипические эффекты мутации и избирательное удаление фенотипов.
Наши результаты имеют важное значение для проверки гипотезы пластичности. Такие тесты часто проводятся путем сравнения популяций предков (2, 5, 6, 19, 37–39).Как правило, исследования выявляют набор признаков, которые разошлись между популяциями, и устанавливают, проявляют ли предковые популяции пластичность в направлении потомков по этим признакам. Однако, хотя согласование между пластичными реакциями и основной осью генетической изменчивости делает дивергенцию, обусловленную пластичностью, более вероятной, оно также затрудняет различение признаков эволюции, обусловленной пластичностью, от эволюционной дивергенции, обусловленной фенотипическими эффектами генетической мутации (т.е., «генетические ограничения»; ссылки 29 и 30). В более общем плане количественные генетические оценки могут дать ограниченное представление о более долгосрочной эволюции (40, 41). Таким образом, различение отчетливых эмпирических следов экологических и генетических предубеждений в развитии в эволюции потребует идентификации соответствующих «нулевых» моделей (38). Сравнение потомков и предков в естественных популяциях в сочетании с теоретическим моделированием и экспериментальными исследованиями эволюции, вероятно, будет необходимо для оценки роли пластичности в адаптивной диверсификации.
Материалы и методы
Поиск литературы и сбор данных.
Мы провели два литературных поиска исследований, содержащих либо ( i ) «G matri *» И «P matri *» И «окружающая среда *», либо ( ii ) «генетическая ковариация» И «фенотип * ковариация» И Environment * ‘в заголовке, аннотации или ключевых словах, чтобы найти исследования, оценивающие матрицы G и P в различных средах. В обоих поисках мы использовали Web of Science и Scopus (дата поиска 9 октября 2017 г.), учитывая, что разные поисковые системы могут давать разные результаты (42).Мы ограничили наши предметные области биологией, науками о растениях, эволюционной биологией, экологией, биологией развития и сельским хозяйством. Кроме того, мы извлекли матрицы корреляции / ковариации P и G из первичных исследований, сопоставленных с предыдущими метаанализами, изучающими экологическую чувствительность G (33, 43), и метаанализом, объединяющим влияние окружающей среды на аддитивную генетическую дисперсию. (32).
Для включения в наш метаанализ первичное исследование должно было ( i ) провести количественный генетический план разведения в двух или более средах, где одна среда считалась репрезентативной для популяции или вида (т.е., не новинка), и одна или несколько сред были новыми, ( ii ) измеряют два или более фенотипических признака, и ( iii ) представляют матрицы корреляции / ковариации G и P для этих признаков или G вместе с чертой означает в обеих средах. Мы определили новую среду как среду, в которой исходная популяция не наблюдалась в недавнем прошлом (более пяти поколений) или где авторы четко заявили о новизне среды в статье.Мы далее классифицировали новую среду как стрессовую, если авторы ( i ) четко заявили, что среда была стрессовой и / или ( ii ) было очевидным снижение выживаемости в новой среде. Мы признаем, что новые условия окружающей среды, которые авторы не считают стрессовыми или где данные о смертности не были предоставлены, также могут считаться стрессовыми во многих случаях, но это различие, тем не менее, потенциально важно, поскольку выборочная смертность может влиять на оценки генетической и фенотипической изменчивости.Соответствующие исследования, признанные соответствующими указанным выше критериям на основании названия и аннотаций, были просмотрены двумя авторами (D.W.A.N. и R.R.). В некоторых исследованиях были представлены только корреляционные матрицы. Эти исследования были включены, если мы могли получить оценки аддитивной генетической дисперсии для набора признаков, чтобы матрицы корреляции могли быть преобразованы в матрицы ковариации (необходимые для получения размеров эффекта, см. Ниже). В случаях, когда была доступна только часть необходимых данных (например, только G , а не P , отсутствующие размеры выборки, отсутствие оценок дисперсии и т. Д.) мы оценили G и P на основе необработанных данных, если они были депонированы авторами в репозитории или если они были напрямую связаны с авторами для получения соответствующей информации. Если эту информацию получить не удалось, мы исключили исследование или, в некоторых случаях, признак из нашего анализа. См. Приложение SI , рис. S1, где представлена полная диаграмма PRISMA. Можно найти все необработанные матричные данные, извлеченные из исследований (44) – 75), код и анализ на https://osf.io/fz7sr/.
Переменные модератора (предиктора).
Мы собрали ряд переменных-модераторов, которые, как мы априори ожидали, объясняют различия в величине эффекта. К ним относятся количество количественно оцененных признаков (т. Е. Общий размер матрицы G ), стрессовая обстановка и план исследования, использованный для оценки матриц G (т. Е. Полусиб или полусиб), которые различают узкие — смысловые и общие оценки аддитивной генетической изменчивости соответственно. Для метаанализов, включающих сравнение G в разных средах, мы также классифицировали величину эффекта как полученную в исследованиях, в которых новая среда считалась стрессовой (т.е., стрессовые или не стрессовые) и включили это в соответствующие анализы. Однако важно отметить, что наша главная цель — не изолировать конкретные средовые предикторы вариации в величине эффекта, а оценить степень, в которой направление пластичности и основная ось аддитивной генетической дисперсии совпадают для популяций, подвергшихся воздействию нового среда.
Матричные размеры эффектов и дисперсия выборки.
Мы были особенно заинтересованы в сравнении матриц G и P в нестандартных и новых средах и между ними, а также в определении того, как стрессовая обстановка, количество признаков и дизайн исследования повлияли на величину изменений в матрицах.Кроме того, нас также интересовала степень, в которой пластические ответы (то есть векторы пластичности) совпадают с генетической изменчивостью ( G ) в необычной и новой среде. Эти вопросы требуют оценок размера эффекта, которые можно использовать для сравнения различных аспектов матриц и их согласования (подробности об используемых мерах размера эффекта обсуждаются ниже). Оценка G и P происходит с различными уровнями точности (76), и любые сравнения как внутри, так и между средами должны пытаться учесть влияние дисперсии выборки, чтобы избежать потенциально ошибочных выводов.
Мы применили гибкий метааналитический подход к оценке размера и дисперсии выборки с помощью матриц, что позволило нам использовать традиционные многоуровневые метааналитические модели, которые более эффективно взвешивают исследования на основе их точности. Учитывая, что не существует единого размера эффекта матрицы, охватывающего все аспекты того, как матрицы могут изменяться, мы использовали серию альтернативных показателей размера эффекта (рис. 6 A – D и подробно описаны ниже) и сгенерировали соответствующие им дисперсии выборки для сравнения G и P матрицы внутри и между средами (рис.6 E ). В то время как мы сравнивали G и P между собой в разных средах, мы сосредоточились на сравнении только G . Это уместно, поскольку G и P обычно были очень похожи (см. Код анализа и результаты на https://osf.io/fz7sr/). Перед моделированием любые неположительно-определенные матрицы (т.е. неположительные собственные значения) были «согнуты», чтобы сделать их положительно-определенными, и мы исключили все строки и столбцы матрицы, в которых генетические или фенотипические отклонения признаков были нулевыми или отрицательными (33).Это гарантировало, что все матрицы, используемые для моделирования, были положительно определенными.
Рис. 6.( A — D ) Примеры различных мер величины эффекта, описывающих изменения в G и P внутри и по всему не новеллу (зеленый — «NN») и новому (оранжевый — «ноябрь»). ») Среды. Хотя и G , и P сравнивались друг с другом и сами по себе, внутри и между средами, мы представляем размеры эффектов, сравнивая матрицы G только для простоты.См. Текст для более подробной информации. ( E ) Схематический пример моделирования Монте-Карло, используемого для создания размеров эффекта и соответствующей дисперсии выборки, описанной в A — D .
Чтобы оценить вариабельность выборки для размеров эффекта на основе матрицы, мы использовали моделирование методом Монте-Карло, где каждая положительно определенная ковариационная матрица (Σ) вместе с соответствующим размером выборки (количество семей или производителей для G и количество особей для P ), был использован для создания 5000 смоделированных наборов данных каждой матрицы признаков из бумаги.Наборы признаков имели ту же структуру ковариации, что и ковариационная матрица, взятая из каждой статьи, но с добавлением случайных вариаций (смоделированных). Предполагалось, что признаки подчиняются многомерному нормальному распределению [MVN∼ (x¯, Σ), где x¯ — вектор средних значений (нули для G и среднее значение каждого признака для P ) с длиной, равной размеру количества признаков] (рис. 6 E ). Сгенерированные таким образом размеры матричных эффектов имеют то преимущество, что дисперсия выборки из разных дизайнов исследований распространяется в разных средах и из разных матриц (т.е., G и P ). Матрицы ковариации, созданные из 5000 смоделированных наборов данных, затем использовались для расчета различных статистических данных на основе матриц (описанных ниже). Из этих распределений мы взяли среднее значение распределения в качестве среднего эффекта для исследования и дисперсию распределения в качестве соответствующей дисперсии выборки в исследованиях.
Чтобы гарантировать, что некоторые черты не будут непропорционально влиять на величину эффекта, мы стандартизировали средние черты вместе с матрицами P и G .Для сравнения количественной генетической эволюционируемости, количественно определяемой аддитивными генетическими вариациями внутри популяций и между популяциями, эти данные следует стандартизировать по среднему значению (15, 77). Поскольку нас также интересовала пластическая реакция, которая измеряется как изменение средних признаков, мы не можем просто разделить на средние признаки для каждого исследования в каждой среде. Поэтому сначала мы вычислили среднее значение средних значений признака в каждом исследовании ( μ ) как μ = x¯NN + x¯Nov2,
, где x¯NN — вектор, содержащий средние значения фенотипических признаков в непривычной среде ( я.е., NN ), а x¯Nov — вектор средних значений фенотипических признаков в новой среде (т.е. Nov ). Затем мы стандартизировали средние (x¯μ) и ковариационные матрицы (Σμ) как x¯μ≡x¯μΣμ≡Σ⊘ (μμ ‘),
, где x¯ — вектор со средними значениями для данного исследования и среды, а Σ представляет собой ковариационную матрицу (матрица G или P ). ⊘ обозначает поэлементное деление. Ниже мы подробно описываем, как мы рассчитывали различные размеры эффекта на основе матриц с использованием стандартизованных средних и ковариационных матриц, но для упрощения записи мы опускаем нижний индекс μ в остальной части текста.
Сравнение изменений стандартизованной дисперсии.
Мы рассчитали изменение общей аддитивной генетической изменчивости многомерного фенотипа (рис. 6 A ). Общее количество вариаций сначала вычислялось для каждой матрицы как сумма всех диагональных элементов Σ (то есть след ковариационной матрицы). Затем мы вычислили стандартизированную разницу в общей дисперсии между не новаторскими и новыми матрицами G (SDVG) как SDV = 2tr (ΣNov) –tr (ΣNN) tr (ΣNov) + tr (ΣNN),
, где tr (ΣNN) и tr (ΣNov) — это следы ковариационных матриц в непривлекательной среде и новой среде, соответственно.Мы не принимали абсолютную разницу в дисперсии, потому что нас интересовало, а также у нас были конкретные прогнозы относительно того, как дисперсия должна измениться в ответ на новые условия окружающей среды. Положительные стандартизованные размеры эффекта указывают на то, что общая дисперсия многомерных фенотипов в новых средах была больше, чем общая дисперсия в не новаторских средах.
Сравнение пропорций отклонения по главным осям
G .Мы также оценили долю вариации в G (PVG_max), которая была вдоль главной оси аддитивной генетической изменчивости (т.е.е., вдоль gmax) для каждой среды (рис. 6 A ). Мы количественно оценили долю общей дисперсии по gmax следующим образом: PVmax = λmaxtr (Σ),
, где λmax — первое собственное значение (то есть величина вариации по первому собственному вектору), а tr (Σ) — след ковариационной матрицы. Учитывая, что PVmax является пропорцией, мы преобразовали ее в логит, чтобы нормализовать ее распределение и ошибку выборки.
Сравнение основных осей генетической и фенотипической изменчивости.
Во-вторых, мы исследовали, как ориентации G и P меняются внутри и между средами, вычислив угол между главными осями изменения (рис.6 В ). Большая ось изменения ковариационной матрицы описывается ее первым собственным вектором. Для сравнения в условиях окружающей среды мы вычислили угол между gmax и pmax (θNN или θNov в зависимости от среды), которые являются первыми собственными векторами G и P соответственно. Для сравнения между средами мы вычислили угол между не нововведением gmax и новым gmax (θG), или угол между не нововведением pmax и новым pmax (θP), в зависимости от того, был ли рассчитан угол для генетической или фенотипической вариации.Угол между двумя первыми собственными векторами был вычислен как θ = 180πcos − 1 (v1_max⋅v2_maxv1_max⋅v1_max⋅v2_max⋅v2_max),
, где v1_max — первый собственный вектор одной из ковариационных матриц, а v2_max — первый собственный вектор ковариации. матрица. В результате этого расчета углы меняются от 0 ° до 180 °, поэтому мы вычли из 180 ° любой угол от 180 ° до 90 °, чтобы получить углы от 0 ° до 90 ° (33). Чтобы нормализовать распределение углов, мы разделили их на 90 и выполнили логит-преобразование.
Оценка краткосрочного эволюционного потенциала пластических реакций.
Наконец, чтобы исследовать краткосрочный эволюционный потенциал по измерениям пластических реакций, мы проверили, совпадает ли вектор пластической реакции — изменение средних значений признаков между не новаторской и новой средой — с матрицами G не новеллы и новое окружение. Сначала мы вычислили вектор Δx¯, описывающий изменение средних значений признаков: Δx¯ = x¯NN − x¯Nov.
Затем мы спроецировали матрицы G как для новой, так и для новой среды на вектор Δx¯, чтобы вычислить величину дисперсии в G в направлении вектора Δx¯ (т. Е. EΔx¯), используя уравнение 1 в исх. 15: eΔx¯ = (Δx¯) ‘GΔx¯ | Δx¯ | 2.
Чтобы оценить, насколько сильно вариация присутствует в направлении пластического отклика по сравнению с максимальной величиной вариации в любом направлении, мы разделили eΔx¯ на первое собственное значение G (λG_max), чтобы вычислить эволюционный потенциал пластичности. (Рисунок.6 C ): πe = eΔx¯λG_max.
Учитывая, что πe является пропорцией, мы преобразовали ее в логит, чтобы нормализовать ее распределение и ошибку выборки. Чтобы проверить, было ли πe значительно больше, чем ожидалось для любого случайного совпадения между пластическим откликом и G , мы также вычислили нулевое ожидание для эволюционного потенциала пластичности (π0): π0 = λ¯GλG_max,
, в котором λ¯ G — это среднее значение всех собственных значений для G . Затем мы вычли логит π0 из логита πe, чтобы оценить разницу между наблюдаемым и ожидаемым эволюционным потенциалом в направлении пластичности (πe − 0): πe − 0 = logit (πe) −logit (π0).
Когда нижняя граница вероятного интервала для пересечения метааналитической модели (как описано ниже) для πe − 0 больше нуля, эволюционный потенциал в направлении пластичности выше, чем мы могли бы ожидать для случайное выравнивание между Δx¯ и gmax. Мы также вычислили угол между Δx¯ и первым собственным вектором G (vG_max) (рис. 6 D ): θe = 180πcos − 1 (Δx¯⋅vG_maxΔx¯⋅Δx¯⋅vG_max⋅vG_max).
В результате этого вычисления углы меняются от 0 ° до 180 °, поэтому мы вычли из 180 ° любой угол от 180 ° до 90 °, чтобы получить углы от 0 ° до 90 °.Чтобы нормализовать распределение углов, мы разделили их на 90, а затем выполнили логит-преобразование.
Мета-анализ.
Мы провели мета-анализ размеров эффекта с использованием многоуровневых метааналитических и метарегрессионных моделей (78) с метафором пакета R (79). Во всех моделях эффекты были взвешены по дисперсии их выборки из моделирования, и мы включили «исследование» как случайный эффект. Для анализа, сравнивающего среды, каждое исследование внесло несколько оценок, поэтому мы включили случайный наклон типа среды в случайный эффект на уровне исследования, чтобы учесть эту независимость.Тем не менее, только случайное пересечение было включено, если отклонение наклона не сходилось в модели. Учитывая, что между исследованием и видами было сильное совпадение, мы не включили случайный эффект на уровне видов или филогению в наши модели, чтобы избежать искажающих оценок (80). Пакет metafor по умолчанию не оценивает остаточную дисперсию, поэтому мы включили случайный эффект на уровне наблюдения во все наши модели. В некоторых исследованиях манипулировали множественными средами, и, учитывая количественный генетический дизайн, часто разделяли семьи в этих средах, это вводит уровень независимости между величинами эффектов (80).Учитывая ограниченный размер выборки, мы не учли этот уровень зависимости; однако мы признаем, что это может привести к завышению количества ошибок типа I.
Мы использовали только метааналитические модели перехвата для количественной оценки общей (It2) и межисследовательской (Ist2) неоднородности в эффектах (81, 82) как Ist2 = σst2σst2 + σe2 + σm2It2 = σst2 + σe2σst2 + σe2 + σm2σm2 = ∑wi (k −1) [(∑wi) 2 − ∑wi2],
, где σm2 — оценка общей изменчивости выборки, полученная из wi, веса или обратной величины дисперсии выборки, и k — общего количества эффектов.σst2 и σe2 — оценки дисперсии между исследованиями и внутри исследований. Ist2 можно интерпретировать как долю вариации в эффектах, возникающих в результате различий между исследованиями, которая включает в себя такие различия, как виды, типы признаков и методологические подходы, которые различаются в разных исследованиях. Напротив, It2 — это общая доля вариации эффектов помимо дисперсии выборки. Если It2 высокое, это говорит о том, что наблюдаемые эффекты не просто результат изменчивости выборки.
Мы запустили модели метарегрессии, чтобы изучить, как наши предсказанные переменные-модераторы объясняют вариации в эффектах.Для каждого размера эффекта мы включили основные эффекты для модераторов, которые, по нашему предположению, могли бы объяснить вариации в эффектах, когда размер выборки позволял. К ним относятся ( i ) количество черт, ( ii ) тип среды (не новаторская и новая — только для анализов, сравнивающих соответствие между вектором пластичности и G ), ( iii ) тип новой среды. (т. е. стрессовые или нестрессовые — только для анализов, сравнивающих G в разных средах) и ( iv ) дизайн разведения (т.э., полусиб или полный сиб). Учитывая ограниченный размер выборки и неравномерную таксономическую выборку, мы ограничили модели оценкой только основных эффектов вышеуказанных модераторов. Модели, сравнивающие G и P в разных средах, были запущены отдельно. Мы рассчитали маргинализованный средний размер эффекта (и доверительные интервалы) для различных подгрупп категориальных предикторов, взяв средневзвешенное значение среднего (и дисперсии выборки) одного уровня в факторе 1 (например, полусиб для фактора дизайна разведения) по оба уровня фактора 2 (т.е., стресс и нестресс для нового фактора окружающей среды). Мы сделали это для всех комбинаций, чтобы получить средние оценки, не зависящие от других категориальных факторов в модели. Мы проверили асимметрию графика воронки (т. Е. Размер эффекта как функцию ошибки выборки), чтобы найти доказательства систематической ошибки публикации. Однако мы не нашли явных доказательств предвзятости публикации в любом из размеров эффекта ( SI Приложение , рис. S2).
Благодарности
Мы благодарим многочисленных авторов за предоставление дополнительных данных и информации.Мы благодарны Шиничи Накагава, Джоэлю Пику, Алистеру Сеньору, Натали Файнер и Стивену Де Лиллю за обсуждение и вдумчивый отзыв по аспектам проекта и конструктивную обратную связь по предыдущим наброскам рукописи. Три анонимных рецензента предоставили очень полезные комментарии к рукописи. Это исследование финансировалось за счет гранта Фонда Джона Темплтона (60501) и стипендии Валленбергской академии (оба — T.U.). D.W.A.N. был поддержан Австралийским исследовательским советом (ARC) Discovery Early Career Research Award (DE150101774) и стипендией проректоров Университета Нового Южного Уэльса.
Сноски
Авторы: D.W.A.N., R.R., and T.U. спланированное исследование; D.W.A.N., R.R. и T.U. проведенное исследование; D.W.A.N. и Р.Р. проанализировали данные; и D.W.A.N., R.R., and T.U. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Размещение данных: Все необработанные матричные данные, код и анализы размещены в репозитории Open Science Framework по адресу https: // osf.io / fz7sr /.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1821066116/-/DCSupplemental.
Изучение фенотипических последствий вариаций тканеспецифической экспрессии генов, выведенных из сводной статистики GWAS
Секция генетической медицины, Чикагский университет, Чикаго, Иллинойс, 60637, США
Альваро Н. Барбейра, Скотт П. Дикинсон , Родриго Бонаццола, Цзямао Чжэн, Каанан П.Шах, Дэн Л. Николае и Хэ Гён Им
Департамент биологии, Университет Лойолы, Чикаго, Чикаго, Иллинойс, 60660, США
Хизер Э. Уиллер
Департамент компьютерных наук, Университет Лойолы Чикаго, Чикаго, Иллинойс, 60660, США
Хизер Э. Уиллер
Комитет по молекулярному метаболизму и питанию, Чикагский университет, Чикаго, Иллинойс, 60637, США
Джейсон М. Торрес
Генетический институт Вандербильта, Университет Вандербильта Медицинский центр, Нашвилл, штат Теннесси, 37232, США
Эрик С.Торстенсон и Нэнси Дж. Кокс
Центр исследовательской информатики, Чикагский университет, Чикаго, Иллинойс, 60615, США
Цинцуни Гарсия
Отдел эпидемиологии, Медицинский факультет, Институт генетики Вандербильта, Медицинский университет Вандербильта Center, Нашвилл, штат Теннесси, 37232, США
Тодд Л. Эдвардс
Отдел психиатрической геномики, Медицинская школа Икана на горе Синай, Нью-Йорк, штат Нью-Йорк, 10029, США
Эли А.Шталь и Лаура М. Хакинс
Департамент генетики и геномики, Медицинская школа Икан в Маунт-Синай, Нью-Йорк, 10029, США
Эли А. Шталь и Лаура М. Хакинс
Институт Броуда Массачусетский технологический институт и Гарвардский университет, Кембридж, Массачусетс, 02142, США
Франсуа Агуэ, Кристин Г. Ардли, Берил Б. Каммингс, Эллен Т. Гельфанд, Гад Гетц, Кейн Хэдли, Роберт Э. Хендсакер, Кэтрин Х. Хуанг , Сева Кашин, Конрад Дж.Карчевски, Монкол Лек, Сяо Ли, Дэниел Г. Макартур, Джаред Л. Недзел, Дуйен Т. Нгуен, Майкл С. Ноубл, Айеллет В. Сегре, Касандра А. Троубридж, Тару Тукиайнен, Мелина Клауснитцер, Лей Хоу, Манолис Келлис, Япин Лю, Бенуа Молини, Юнджин Парк, Никола Дж. Ринальди, Ли Ван и Николас Ван Виттенберги
Отдел аналитической и трансляционной генетики, Массачусетская больница общего профиля, Бостон, Массачусетс, 02114, США
Берил Б. Каммингс, Конрад Дж. Карчевский, Монкол Лек, Даниэль Г.МакАртур и Тару Тукиайнен
Онкологический центр Массачусетской больницы общего профиля, Отделение патологии, Массачусетская больница общего профиля, Бостон, Массачусетс, 02114, США
Гад Гетц
Департамент генетики, Гарвардская медицинская школа 02114, Массачусетс, Массачусетс , США
Роберт Э. Хэндсакер и Сева Кашин
Департамент генетики, Стэнфордский университет, Стэнфорд, Калифорния, , США
Натан С. Абелл, Джо Р. Дэвис, Лор Фрезар, Майкл Дж.Глудеманс, Боксанг Лю, Стивен Б. Монтгомери, Захари Заппала, Джоан Чан, Руйки Цзянь, Лихуа Цзян, Джин Билли Ли, Цинь Ли, Сяо Ли, Джессика Линь, Шин Лин, Сандра Линдер, Стивен Б. Монтгомери, Кевин С. Смит , Майкл П. Снайдер, Хуа Тан, Мэн Ван и Руи Чжан
Кафедра патологии, Стэнфордский университет, Стэнфорд, Калифорния, , США
Натан С. Абелл, Брунильда Баллиу, Джо Р. Дэвис, Лор Фрезард, Майкл Дж. Глодеманс, Синь Ли, Боксян Лю, Стивен Б. Монтгомери, Эмили К.Цанг, Захари Заппала, Сандра Линдер, Стивен Б. Монтгомери, Кевин С. Смит и Эмили К. Цанг
Кафедра клинической биохимии и фармакологии, факультет медицинских наук, Университет Бен-Гуриона в Негеве, Беэр-Шева, 84105, Израиль
Рут Баршир, Омер Баша и Эсти Йегер-Лотем
Департамент компьютерных наук, Университет Джона Хопкинса, Балтимор, Мэриленд, 21218, США
Алексис Батл, Фархан Н. Дамани, Юнгил Ким, Принси Парсана и Ашис Саха
Центр геномного регулирования (CRG), Барселонский институт науки и технологий, 08003, Барселона, Испания
Гириш К.Bogu, Diego Garrido-Martín, Roderic Guigo, Jean Monlong, Manuel Muñoz-Aguirre, Panagiotis Papasaikas, Ferran Reverter & Reza Sodaei
Universitat Pompeu Fabra (UPF), 08002 Bogirees, Барселона, Испания
Гарридо-Мартин, Родерик Гиго, Жан Монлонг, Мануэль Муньос-Агирре, Панайотис Папасайкас, Ферран Ревертер и Реза СодаиДепартамент генетической медицины и развития, Медицинская школа Женевского университета, 1211, Женева, Швейцария
Эндрю Браун, Оливье Делано, Эммануил Т.Дермитзакис, Седрик Ховальд, Халит Онген и Николаос Панусис
Институт генетики и геномики в Женеве (iG3), Женевский университет, 1211, Женева, Швейцария
Эндрю Браун, Оливье Делано, Эммануил Т. Дермитзакис, Седрик Ховальд Халит Онген и Николаос Панусис
Швейцарский институт биоинформатики, 1211, Женева, Швейцария
Эндрю Браун, Оливье Делано, Эммануил Т. Дермитзакис, Седрик Ховальд, Халит Онген и Николаос Панусис
of Medicine, Пенсильванский университет, Филадельфия, Пенсильвания, 19104, США
Кристофер Д.Brown & YoSon Park
New York Genome Center, New York, NY, 10013, USA
Стефан Э. Кастель, Сара Ким-Хельмут, Туули Лаппалайнен и Педжман Мохаммади
Департамент системной биологии, Медицинский университет Колумбийского университета Center, New York, NY, 10032, USA
Стефан Э. Кастель, Сара Ким-Хельмут, Туули Лаппалайнен и Пейман Мохаммади
Департамент здравоохранения, Чикагский университет, Чикаго, Иллинойс, 60637, США
Лин С.Чен, Лин С. Чен, Кэтрин Деманелис, Фарзана Жасмин, Мухаммад Г. Кибрия и Брэндон Л. Пирс
Институт генома Макдоннелла, Школа медицины Вашингтонского университета, Сент-Луис, Миссури, 63108, США
Колби Чианг, Ира М. Холл и Александра Дж. Скотт
Отделение генетики Медицинского факультета Вашингтонского университета, Сент-Луис, Миссури, 63108, США
Дональд Ф. Конрад и Ира М. Холл
Отделение патологии И иммунология, Медицинский факультет Вашингтонского университета, Св.Луис, Миссури, 63108, США
Дональд Ф. Конрад
Департамент компьютерных наук, Центр статистики и машинного обучения, Принстонский университет, Принстон, Нью-Джерси, 08540, США
Барбара Э. Энгельхардт
Департамент of Computer Science, Калифорнийский университет, Лос-Анджелес, Калифорния, , США
Элеазар Эскин, Фархад Хормоздиари, Ын Йонг Канг и Сергей Мангул
Департамент генетики человека, Калифорнийский университет, Лос-Анджелес, Калифорния, США
Элеазар Эскин
Instituto de Investigação e Inovação em Saúde (i3S), Universidade do Porto, 4200-135, Порту, Португалия
Pedro G.Феррейра
Институт молекулярной патологии и иммунологии (IPATIMUP), Университет Порту, 4200-625, Порту, Португалия
Педро Г. Феррейра
Отдел генетической медицины, Медицинский центр, Медицинский центр Университета Вандербильта, Нашвилл, Теннесси, 37232, США
Эрик Р. Гамазон
Отдел клинической эпидемиологии, биостатистики и биоинформатики, Академический медицинский центр, Амстердамский университет, 1105 AZ, Амстердам, Нидерланды
Эрик Р.Gamazon
Департамент психиатрии, Академический медицинский центр, Амстердамский университет, 1105 AZ, Амстердам, Нидерланды
Эрик Р. Гамазон
Институт Льюиса Сиглера, Принстонский университет, Принстон, штат Нью-Джерси, 08540, США
Ariel DH Gewirtz, Brian Jo & Joshua M. Akey
Департамент операционных исследований и финансового инжиниринга, Принстонский университет, Принстон, Нью-Джерси, 08540, США
Genna Gliner
Программа биомедицинской информатики, Стэнфордский университет, Стэнфорд, CA, , USA
Майкл Дж.Глудеманс, Эмили К. Цанг и Эмили К. Цанг
Institut Hospital del Mar d’Investigacions Mèdiques (IMIM), 08003, Барселона, Испания
Родерик Гиго
Департамент медицины, Школа медицины Вашингтонского университета, Сент-Луис, Миссури, 63108, США
Ира М. Холл
Кафедра медицины конвергенции, Медицинский колледж Ульсанского университета, Медицинский центр Асан, Сеул, 138-736, Южная Корея
Бум Хан
Департамент биомедицинской инженерии, Университет Джона Хопкинса, Балтимор, Мэриленд, 21218, США
Юань Хе, Бенджамин Дж.Стробер и Эндрю П. Файнберг
Департамент биостатистики, Школа общественного здравоохранения Mailman, Колумбийский университет, Нью-Йорк, штат Нью-Йорк, 10032, США
Ген Ли
Департамент биологии Стэнфордского университета, Стэнфорд, Калифорния, , USA
Boxiang Liu
Wellcome Trust Center for Human Genetics, Медицинский факультет Nuffield, Оксфордский университет, Оксфорд, OX3 7BN, UK
Mark I. McCarthy, Anne W. Ndungu, Anthony J.Пейн и Мартин ван де Бунт
Оксфордский центр диабета, эндокринологии и метаболизма, Оксфордский университет, Больница Черчилля, Оксфорд, OX3 7LE, Великобритания
Марк И. Маккарти и Мартин ван де Бунт
Оксфорд Никал Исследовательский центр, больница Черчилля, Оксфорд, OX3 7LJ, Великобритания
Марк И. Маккарти
Программа выпускников по вычислительной биологии и биоинформатике, Университет Дьюка, Дарем, Северная Каролина, 27708, США
Ян К.МакДауэлл
Кафедра генетики человека, Университет Макгилла, Монреаль, Квебек, h4A 0G1, Канада
Жан Монлонг
Departament d’Estadística i Investigació Operativa, Universitat Politècnica2000, Испания, 9000, Испания, 9000, Испания -Aguirre
Департамент статистики и исследований операций, Университет Северной Каролины, Чапел-Хилл, Северная Каролина, 27599, США
Эндрю Б. Нобель и Джон Дж. Палович
Департамент биостатистики Университета Северной Каролины, Чапел-Хилл, Северная Каролина, 27599, США
Эндрю Б.Nobel
Секция генетической медицины, Медицинский факультет, Чикагский университет, Чикаго, Иллинойс, 60637, США
Мериткселл Олива, Барбара Э. Стрейнджер, Мариан С. Фернандо, Кэролайн Линке, Мериткселл Олива, Эндрю Скол, Барбара Э. Стрейнджер и Фан Ву
Институт геномики и системной биологии, Чикагский университет, Чикаго, Иллинойс, 60637, США
Мериткселл Олива, Барбара Э. Стрейнджер, Мариан С. Фернандо, Кэролайн Линке, Мериткселл Олива , Эндрю Скол, Барбара Э.Stranger & Fan Wu
Департамент биостатистики, Онкологический центр Андерсона Университета Техаса, Хьюстон, Техас, 77030, США
Кристин Б. Петерсон
Computational Sciences, Pfizer Inc, Кембридж, Массачусетс, 02139, США
Джи Куан и Хуалин С. Си
Барселонский университет, 08028, Барселона, Испания
Ферран Ревертер
Департамент биомедицинских данных, Стэнфордский университет, Стэнфорд, Калифорния, , США
Сабатти
Статистический факультет Стэнфордского университета, Стэнфорд, Калифорния, , США
Кьяра Сабатти
Институт биофизики Карлоса Шагаса Филью (IBCCF), Федеральный университет Рио-де-Жанейро (UFRJ), 212, 212, де Жанейро, Бразилия
Майкл Саммет
Кафедра психиатрии, Университет штата Юта, Солт-Лейк-Сити, Юта, 84108, США
Андрей А.Шабалин
Департамент статистики Чикагского университета, Чикаго, Иллинойс, 60637, США
Мэтью Стивенс
Департамент генетики человека, Чикагский университет, Чикаго, Иллинойс, 60637, США
Мэтью Стивенс , Сара Урбут и Гао Ван
Центр интенсивных исследований данных, Чикагский университет, Чикаго, Иллинойс, 60637, США
Барбара Э. Стрейнджер, Эндрю Скол и Барбара Э. Стрейнджер
Департамент психиатрии и Биоповеденческие науки, Калифорнийский университет, Лос-Анджелес, Калифорния, , США
Джэ Хун Сул
Кафедра биостатистики, Мичиганский университет, Анн-Арбор, Мичиган, 48109, США
Сяокван Вен
Bioinformics Research Центр и департаменты статистики и биологических наук, Государственный университет Северной Каролины, Роли, Северная Каролина, 27695, США
Фред А.Wright & Yi-Hui Zhou
Национальный институт биотехнологии в Негеве, Беэр-Шева, 84105, Израиль
Эсти Йегер-Лотем
Европейская лаборатория молекулярной биологии, 69117, Гейдельберг, Германия
Джудит Заугг
Департамент экологии и эволюционной биологии Принстонского университета, Принстон, Нью-Джерси, 08540, США
Джошуа М. Эйки
Институт биомедицинских наук Альтиуса, Сиэтл, Вашингтон, 98121, США
Дэниел Бейтс, Морган Дигель, Джессика Хэлоу, Эрик Хоген, Одра Джонсон, Раджиндер Каул, Кристен Ли, Джемма Нельсон, Фиденсио Дж.Нери, Ричард Сандстром и Джон Стаматояннопулос
Медицинский центр Бет Исраэль, Гарвардская медицинская школа, Бостон, Массачусетс, 02215, США
Мелина Клауснитцер
Университет Хоэнхайм2, 70599, Германия
, ШтутгартИнститут рака Хантсмана, Департамент наук о здоровье населения, Университет Юты, Солт-Лейк-Сити, Юта, 84112, США
Дженнифер А. Доэрти
Центр эпигенетики, Школа медицины Университета Джона Хопкинса, Балтимор, Мэриленд , 21205, США
Эндрю П.Файнберг, Каспер Д. Хансен и Линдси Ф. Риццарди
Департамент медицины, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205, США
Эндрю П. Фейнберг
Департамент психического здоровья, Джонс Хопкинс Университетская школа общественного здравоохранения, Балтимор, Мэриленд, 21205, США
Эндрю П. Фейнберг
Институт генетической медицины МакКусика-Натанса, Медицинская школа Джона Хопкинса, Балтимор, Мэриленд, 21205, США
Каспер Д.Хансен
Департамент биостатистики, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205, США
Каспер Д. Хансен и Питер Ф. Хики
Лаборатория компьютерных наук и искусственного интеллекта, Массачусетский технологический институт, Кембридж, Массачусетс, Массачусетс , 02139, USA
Lei Hou, Manolis Kellis, Yaping Liu, Yongjin Park и Nicola J. Rinaldi
Департамент медицины, Вашингтонский университет, Сиэтл, Вашингтон, 98195, США
Джессика Лин и Джон Стаматояннопулос
Отделение кардиологии Вашингтонского университета, Сиэтл, Вашингтон, 98195, США
Шин Лин
Институт системной генетики, Медицинский центр Лангоне Нью-Йоркского университета, Нью-Йорк, Нью-Йорк, 10016, США
Мэтью Т.Маурано
Департамент геномных наук, Вашингтонский университет, Сиэтл, Вашингтон, 98195, США
Джон Стаматояннопулос
Управление стратегической координации, Отдел координации программ, планирования и стратегических инициатив, Офис директора, NIH , Rockville, MD, 20852, США
Консепсьон Р. Ньеррас
Отделение биорепозиториев и исследований биологических образцов, Отдел лечения и диагностики рака, Национальный институт рака, Бетесда, Мэриленд, 20892, США
Филип А.Брантон, Латарша Дж. Каритерс, Пинг Гуан, Хелен М. Мур, Абхи Рао и Джимми Б. Вотут
Национальный институт стоматологических и черепно-лицевых исследований, Бетесда, Мэриленд, 20892, США
Латарша Дж. Каритерс
Отдел геномной медицины, Национальный институт исследования генома человека, Роквилл, Мэриленд, 20852, США
Сара Э. Гулд, Николь К. Локкарт, Кейси Мартин, Джеффри П. Струэвинг и Симона Вольпи
Отдел неврологии и фундаментальной науки Наука о поведении, Национальный институт психического здоровья, NIH, Bethesda, MD, 20892, США
Анджен М.Аддингтон и Сьюзан Э. Кестер
Отдел неврологии и поведения, Национальный институт злоупотребления наркотиками, NIH, Бетесда, Мэриленд, 20892, США
А. Роджер Литтл
Вашингтонское региональное сообщество трансплантологов, Фоллс-Черч, Вирджиния , 22003, USA
Lori E. Brigham
Gift of Life Donor Program, Филадельфия, Пенсильвания, 19103, США
Ричард Хас
LifeGift, Хьюстон, Техас, 77055, США
Кевин Маркус Хантер Майер и Брайан Роу
Центр восстановления и обучения органов, Питтсбург, Пенсильвания, 15238, США
Кристофер Джонс и Джозеф Уиллер
LifeNet Health, Вирджиния-Бич, Вирджиния, 23453, США
Марк Джонсон, Гэри Уолтерс и Майкл Вашингтон
National Disease Research Interchange, Филадельфия, Пенсильвания, 19103, США
Джин Копен, Уильям Ф.Лейнвебер, Джон Т. Лонсдейл, Алиса Макдональд, Бернадетт Местичелли, Майкл Сальваторе, Сабур Шад и Джеффри А. Томас
Unyts, Buffalo, NY, 14203, США
Джейсон Бридж и Марк Миклос
, Roswell Park Cancer Institute, Buffalo, NY, 14263, USA
Барбара А. Фостер, Брайан М. Гиллард, Эллен Карасик, Рахна Кумар и Майкл Т. Мозер
Исследовательский институт Ван Андел, Гранд-Рапидс, Мичиган, 49503 , США
Скотт Д.Джуэлл, Роберт Г. Монтрой, Дэниел К. Рорер и Дана Р. Вэлли
Brain Endowment Bank, Медицинская школа Миллера, Университет Майами, Майами, Флорида, 33136, США
Дэвид А. Дэвис и Дебора К. Маш
Национальный институт аллергии и инфекционных заболеваний, NIH, Роквилл, Мэриленд, 20852, США
Анита Х. Ундейл
Группа по исследованию биопрепаратов, Управление клинических исследований, Leidos Biomedical Research, Inc., Роквилл, Мэриленд , 20852, США
Анна М.Смит, Дэвид Э. Табор, Нэнси В. Рош, Джеффри А. Маклин, Негин Ватанян, Карна Л. Робинсон, Лесли Собин, Кимберли М. Валентино, Ликун Ци, Стивен Хантер, Пушпа Харихаран, Шилпи Сингх, Ки Сунг Ум, Такунда Матос и Мария М. Томашевски
Leidos Biomedical Research, Inc, Frederick, MD, 21701, USA
Mary E. Barcus
Temple University, Philadelphia, PA, 19122, USA
Laura K. Barker, Лаура А. Симинофф и Хизер М. Трейно
Департамент поведения и политики в области здравоохранения, Школа медицины, Университет Содружества Вирджинии, Ричмонд, штат Вирджиния, 23298, США
Maghboeba Mosavel
Европейская лаборатория молекулярной биологии, Европейская лаборатория биоинформатики Institute, Hinxton, CB10 1SD, UK
Paul Flicek, Thomas Juettemann, Magali Ruffier, Dan Sheppard, Kieron Taylor, Stephen J.Треванион и Даниэль Р. Зербино
Институт геномики UCSC, Калифорнийский университет Санта-Крус, Санта-Крус, Калифорния,, США
Брайан Крафт, Мэри Голдман, Максимилиан Хойсслер, У. Джеймс Кент, Кристофер М. Ли, Бенедикт Патен, Кейт Р. Розенблум, Джон Вивиан и Джинчун Чжу
ANB участвовал в разработке программного обеспечения S-PrediXcan; выполненный S-PredixCan запускается по признакам GWAS; разработан механизм для сравнения PrediXcan и S-PrediXcan в моделируемых, клеточных фенотипах и фенотипах WTCCC; запускал COLOC и SMR; разработан gene2pheno.база данных организации и веб-панель управления; внесла свой вклад в основной текст, приложение, рисунки и анализы. S.P.D. провел обучение модели GTEx; запустил GERA GWAS; способствовал основному тексту. J.M.T. способствовал основному тексту. J.Z. внесла свой вклад в цифры и ресурс predictdb.org. СТАНДАРТНОЕ ВОСТОЧНОЕ ВРЕМЯ. внесла свой вклад в программное обеспечение S-PrediXcan. H.E.W. способствовал основному тексту и анализу. К.П.С. запустил PrediXcan на данных WTCCC и внес свой вклад в анализ. Р.Б. внес свой вклад в основной текст и рисунки. T.G. проведено вменение генотипов GERA.T.L.E. способствовал анализу. L.M.H. способствовал основному тексту. E.A.S. способствовал основному тексту. D.L.N. способствовал основному тексту и анализу. Нью-Джерси способствовал анализу. H.K.I. разработал метод, руководил проектом, выполнил анализ, внес вклад в основной текст, приложение и рисунки. Авторы консорциума GTEx внесли свой вклад в сбор, сбор и обработку данных исследований GTEx, используемых для обучения моделей прогнозирования транскриптомов и запуска COLOC и SMR.
Молекулярная основа социально опосредованной фенотипической пластичности осы эусоциальной бумаги
Хадфилд, Дж. Д., Наталл, А., Осорио, Д. и Оуэнс, IPF Тестирование фенотипического гамбита: фенотипические, генетические и экологические корреляции цвета . J. Evol. Биол. 20 , 549–557 (2007).
CAS PubMed Статья PubMed Central Google Scholar
Rittschof, C.К. и Робинсон, Дж. Э. Геномика: продвижение поведенческой экологии за пределы фенотипического гамбита. Аним. Behav. 92 , 263–270 (2014).
PubMed PubMed Central Статья Google Scholar
Рубин, Х. Фенотипический гамбит: давление отбора и методология ESS в эволюционной теории игр. Biol. Филос. 31 , 551–569 (2016).
Артикул Google Scholar
Реале Д., Дингеманс Н. Дж., Казем А. Дж. И Райт Дж. Эволюционные и экологические подходы к изучению личности. Philos. Пер. R. Soc. Б. 365 , 3947–3946 (2010).
Артикул Google Scholar
Чепмен, Б. Б., Брёнмарк, К., Нильссон, Дж. А.. И Ханссон, Л. А. Экология и эволюция частичной миграции. Ойкос 120 , 1764–1775 (2011).
Артикул Google Scholar
Фаулер-Финн, К. Д. и Родригес, Р. Л. Эволюция опосредованной опытом пластичности в предпочтениях партнера. J. Evol. Биол. 25 , 1855–1863 (2012).
PubMed Статья PubMed Central Google Scholar
Хейес, К. Блэкбоксинг: стратегии социального обучения и культурная эволюция. Philos. Пер. R. Soc. В 371 , 20150369 (2016).
Артикул Google Scholar
Horiguchi, R. et al. Характеристика экспрессии фактора гонадной сомы во время смены пола у протогинного губана, Halichoeres trimaculatus. Dev. Дин. 242 , 388–399 (2013).
CAS PubMed Статья PubMed Central Google Scholar
Casas, L. et al. Смена пола у рыб-клоунов: молекулярные выводы из анализа транскриптома. Sci. Реп. 6 , 35461 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Тодд, Э. В. и др. Стресс, новые половые гены и эпигенетическое перепрограммирование организуют социально контролируемую смену пола. Sci. Adv. 5 , eaaw7006 (2019).
ADS CAS PubMed PubMed Central Статья Google Scholar
Каллен, Д.A. et al. От молекул к управлению: механизмы и последствия полифенизма фазы саранчи. Adv. Insect Physiol. 53 , 167–285 (2017).
Артикул Google Scholar
Ло, Н., Симпсон, С. Дж. И Меч, Г. А. Эпигенетика и пластичность развития у ортоптероидных насекомых. Curr. Opin. Insect Sci. 25 , 25–34 (2018).
PubMed Статья PubMed Central Google Scholar
Simola, D. F. et al. Эпигенетическое (ре) программирование кастоспецифического поведения муравья Camponotus floridanus . Наука 351 , aac6633 (2016).
PubMed PubMed Central Статья CAS Google Scholar
Gospocic, J. et al. Нейропептид коразонин контролирует социальное поведение и кастовую принадлежность муравьев. Ячейка 170 , 748–759 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Либбрехт, Р., Оксли, П. Р. и Кронауэр, Д. Дж. Транскриптомика мозга клональных налетчиков-налетчиков определяет молекулярные механизмы-кандидаты для репродуктивного разделения труда. BMC Biol. 16 , 89 (2018).
PubMed PubMed Central Статья CAS Google Scholar
Rehan, S. M. et al. Консервативные гены лежат в основе фенотипической пластичности у начинающей социальной пчелы. Genome Biol. Evol. 10 , 2749–2758 (2018).
CAS PubMed PubMed Central Статья Google Scholar
Шелл, У. А. и Рехан, С. М. Поведенческие и генетические механизмы социальной эволюции: идеи изначально и факультативно социальных пчел. Apidologie 49 , 13–30 (2018).
CAS Статья Google Scholar
Швандер Т., Ло Н., Бикман М., Олдройд Б. П. и Келлер Л. Природа против воспитания в кастовой дифференциации социальных насекомых. Trends Ecol. Evol. 25 , 275–282 (2010).
PubMed Статья PubMed Central Google Scholar
Strassmann, J. E. et al. Цена потери матки у социальной осы Polistes dominulus (Hymenoptera: Vespidae). Дж. Канс. Энтомол. Soc. 77 , 343–355 (2004).
Артикул Google Scholar
Тиббетс, Э. А. и Хуанг, З. Ю. Гипотеза о вызове у насекомого: уровень ювенильного гормона увеличивается во время репродуктивного конфликта после потери матки у ос Polistes . Am. Nat. 176 , 123–130 (2010).
PubMed Статья PubMed Central Google Scholar
Чандра, В.и другие. Социальная регуляция передачи сигналов инсулина и эволюция эусоциальности у муравьев. Наука 361 , 398–402 (2018).
ADS CAS PubMed PubMed Central Статья Google Scholar
Старкс, П. Т., Туриллацци, С. и Вест-Эберхард, М. Дж. Полистес бумажные осы: появление модельного рода. Ann. Zool. Фенн. 43 , 385–386 (2006).
Google Scholar
Джандт, Дж. М., Тиббетс, Э. А. и Тот, А. Л. Polistes бумажные осы: модельный род для изучения иерархий социального доминирования. Insectes Soc. 61 , 11–27 (2014).
Артикул Google Scholar
Standage, D. S. et al. Секвенирование генома, транскриптома и метилома примитивной эусоциальной осы обнаруживает значительно сниженную систему метилирования ДНК у социальных насекомых. Мол.Ecol. 25 , 1769–1784 (2016).
CAS PubMed Статья PubMed Central Google Scholar
Geffre, A.C. et al. Транскриптомика расширенного фенотипа: паразитарные манипуляции с социальным поведением осы изменяют экспрессию связанных с кастом генов. Proc. R. Soc. В 284 , 20170029 (2017).
PubMed Статья CAS PubMed Central Google Scholar
Манфредини, Ф., Браун, М. Дж. И Тот, А. Л. Гены-кандидаты для сотрудничества и агрессии у социальной осы Polistes dominula . J. Comp. Physiol. А 204 , 449–463 (2018).
CAS Статья Google Scholar
Парди, Л. Порядок доминирования в Полистесе ос. Physiol. Zool. 21 , 1–13 (1948).
CAS PubMed Статья PubMed Central Google Scholar
Tsuji, K. & Tsuji, N. Почему иерархия доминирования у социальных насекомых связана с возрастом? Гипотеза относительного долголетия. Behav. Ecol. Sociobiol. 58 , 517–526 (2005).
Артикул Google Scholar
Тейлор, Б.А., Чини, А., Черво, Р., Ройтер, М., и Самнер, С. Конфликт наследования королевы у бумажной осы Polistes dominula смягчается возрастным условием. Behav. Экол . 31 , araa045 (2020).
Segal, N.H. et al. Классификация и прогнозирование подтипов саркомы мягких тканей у взрослых по функциональной геномике. Am. J. Pathol. 163 , 691–700 (2003).
CAS PubMed PubMed Central Статья Google Scholar
Abeel, T., Helleputte, T., Van de Peer, Y., Dupont, P. & Saeys, Y. Надежная идентификация биомаркеров для диагностики рака с помощью методов выбора ансамбля признаков. Биоинформатика 26 , 392–398 (2010).
CAS PubMed Статья PubMed Central Google Scholar
Huang, S. et al. Применение машинного обучения опорных векторов (SVM) в геномике рака. Cancer Genom. Proteom. 15 , 41–51 (2018).
CAS Google Scholar
Чакраварти, П., Коззи, Г., Озгул А. и Аминиан К. Новый биомеханический подход к распознаванию поведения животных с использованием акселерометров. Methods Ecol. Evol. 10 , 802–814 (2019).
Артикул Google Scholar
Ноубл, В. С. Что такое машина опорных векторов? Нат. Biotechnol. 24 , 1565–1567 (2006).
CAS PubMed Статья PubMed Central Google Scholar
Toth, A. L. et al. Общие гены, связанные с агрессией, а не с химической связью, связаны с репродуктивным доминированием у бумажных ос ( Polistes metricus ). BMC Genomics 15 , 75 (2014).
PubMed PubMed Central Статья CAS Google Scholar
Patalano, S. et al. Молекулярные сигнатуры пластических фенотипов у двух видов эусоциальных насекомых с простыми сообществами. Proc. Natl. Акад. Sci. США 112 , 13970–13975 (2015).
ADS CAS PubMed Статья PubMed Central Google Scholar
Мияно, С. Размножение рабочих и родственное поведение в сиротских колониях японской бумажной осы, Polistes jadwigae (Hymenoptera, Vespidae). J. Ethol. 9 , 135–146 (1991).
Артикул Google Scholar
Даппорто Л., Следж Ф. М. и Туриллацци С. Динамика химических профилей кутикулы Polistes dominulus рабочих в осиротевших гнездах (Hymenoptera, Vespidae). J. Insect Physiol. 51 , 969–973 (2005).
CAS PubMed Статья PubMed Central Google Scholar
Wojciechowski, M. et al. Фенотипически различные касты самок медоносных пчел определяются альтернативными состояниями хроматина во время личиночного развития. Genome Res. 28 , 1532–1542 (2018).
CAS PubMed PubMed Central Статья Google Scholar
Дункан, Э. Дж., Лиск, М. П. и Дирден, П. К. Архитектура генома способствует фенотипической пластичности у медоносной пчелы ( Apis mellifera ). Мол. Биол. Evol. 37 , 1964–1978 (2020).
CAS PubMed PubMed Central Статья Google Scholar
Patalano, S. et al. Специализация и пластичность примитивного социального насекомого. Препринт на https://www.biorxiv.org/content/10.1101/2020.03.31.007492v1 (2020).
Matson, C. K. et al. DMRT1 предотвращает репрограммирование самок в послеродовых семенниках млекопитающих. Природа 476 , 101–104 (2011).
CAS PubMed PubMed Central Статья Google Scholar
Йошимура, Х.& Yamada, Y. Y. Определение кастовой судьбы в первую очередь происходит после появления взрослой особи у примитивно эусоциальной бумажной осы: значение фотопериода во время взрослой стадии. Sci. Nat. 105 , 15 (2018).
Артикул CAS Google Scholar
Белл А. М. и Робинсон Г. Е. Поведение и динамический геном. Наука 332 , 1161–1162 (2011).
ADS CAS PubMed PubMed Central Статья Google Scholar
Rittschof, C.C. et al. Нейромолекулярные реакции на социальный вызов: общие механизмы у мышей, колюшки и медоносной пчелы. Proc. Natl Acad. Sci. США 111 , 17929–17934 (2014).
ADS CAS PubMed Статья PubMed Central Google Scholar
Monnin, T., Cini, A., Lecat, V., Fédérici, P. & Doums, C. Фактического конфликта по поводу наследования колонии нет, несмотря на высокий потенциал конфликта в социальной осе Polistes dominulus . Proc. R. Soc. B 276 , 1593–1601 (2009).
PubMed Статья PubMed Central Google Scholar
Тот, А. Л. и Рехан, С. М. Молекулярная эволюция социальности насекомых: перспектива эко-эволюции. Анну. Преподобный Энтомол. 62 , 419–442 (2017).
CAS PubMed Статья PubMed Central Google Scholar
Гомульский, Л. М. и др. Транскриптомный анализ полового созревания и спаривания у средиземноморской плодовой мухи, Ceratitis capitata . PLoS ONE 7 , e30857 (2012).
Чжоу С., Маккей Т. Ф. и Анхольт Р. Р. Транскрипционные и эпигенетические реакции на спаривание и старение у Drosophila melanogaster . BMC Genom. 15 , 927 (2014).
Артикул CAS Google Scholar
Manfredini, F. et al. Нейрогеномные признаки успехов и неудач в переходах жизненного цикла одного из основных насекомых-опылителей. Genome Biol. Evol. 9 , 3059–3072 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Робинсон, М. Д., Маккарти, Д. Дж. И Смит, Г. К. edgeR: пакет Bioconductor для анализа дифференциальной экспрессии цифровых данных экспрессии генов. Биоинформатика 26 , 139–140 (2010).
CAS Статья Google Scholar
Лав М., Андерс С. и Хубер У. Дифференциальный анализ данных подсчета — пакет DESeq2. Genome Biol. 15 , 10–1186 (2014).
Артикул CAS Google Scholar
Таразона, С., Гарсия, Ф., Феррер, А., Допазо, Дж.& Conesa, A. NOIseq: метод дифференциальной экспрессии RNA-seq, устойчивый к ошибкам глубины секвенирования. EMBnet. J. 17 , 18–19 (2011).
Артикул Google Scholar
Далман, М. Р., Дитер, А., Нимишакави, Г. и Дуан, З. Х. Изменение складывания и отсечки значений p значительно изменяют интерпретацию микрочипов. BMC Bioinform. 13 , 11 (2012).
Артикул Google Scholar
Jung, K., Friede, T. & Beißbarth, T. Сообщение FDR об аналогичных доверительных интервалах для логарифмического изменения дифференциально экспрессируемых генов. BMC Bioinform. 1 , 288 (2011).
Артикул Google Scholar
Шнайдер, К. А., Расбанд, В. С. и Элисейри, К. В. NIH Image to ImageJ: 25 лет анализа изображений. Нат. Методы 9 , 671–675 (2012).
CAS PubMed PubMed Central Статья Google Scholar
Чини, А., Мекончелли, С. и Черво, Р. Индексы яичников как индикаторы репродуктивного вклада и яйцекладки у социальных насекомых: сравнение методов. Insectes Soc. 60 , 393–402 (2013).
Артикул Google Scholar
Friard, O. & Gamba, M. BORIS: бесплатное универсальное программное обеспечение с открытым исходным кодом для регистрации событий для кодирования видео / аудио и наблюдений в реальном времени. Methods Ecol.Evol. 7 , 1325–1330 (2016).
Артикул Google Scholar
Эло, А. Э. Рейтинг шахматистов прошлого и настоящего. (Arco Pub, Нью-Йорк, 1978).
Google Scholar
Neumann, C. et al. Оценка иерархий доминирования: подтверждение и преимущества прогрессивной оценки с помощью Elo-rating. Аним. Behav. 82 , 911–921 (2011).
Артикул Google Scholar
Копылова, Э., Ноэ, Л. и Тузе, Х. SortMeRNA: быстрая и точная фильтрация рибосомных РНК в метатранскриптомических данных. Биоинформатика 28 , 3211–3217 (2012).
CAS PubMed Статья PubMed Central Google Scholar
Bolger, A. M., Lohse, M. и Usadel, B. Trimmomatic: гибкий триммер для данных последовательности Illumina. Биоинформатика 30 , 2114–2120 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Dobin, A. et al. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика 29 , 15–21 (2013).
CAS Статья Google Scholar
Kovaka, S. et al. Сборка транскриптома из выравниваний длинных последовательностей РНК с помощью StringTie2. Genome Biol. 20 , 1–13 (2019).
Артикул CAS Google Scholar
Andrews, S. FastQC: инструмент контроля качества для данных последовательности с высокой пропускной способностью. Доступно в Интернете по адресу: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (2010).
R Основная команда. R: Язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. Доступно в Интернете по адресу: https: // www.R-project.org/ (2020).
Meyer, D., Dimitriadou, E., Hornik, K., Weingessel, A. & Leisch, F. e1071: Различные функции Департамента статистики, группа теории вероятностей (ранее: E1071), TU Wien. Пакет R версии 1.6-8. https://CRAN.R-project.org/package=e1071. (2017).
Langfelder, P. & Horvath, S. WGCNA: пакет R для взвешенного корреляционного сетевого анализа. BMC Bioinform. 9 , 559 (2008).
Артикул CAS Google Scholar
Эммс, Д. М. и Келли, С. OrthoFinder: вывод филогенетической ортологии для сравнительной геномики. Genome Biol. 20 , 1–14 (2019).
Артикул Google Scholar
Smedley, D. et al. Портал сообщества BioMart: инновационная альтернатива крупным централизованным хранилищам данных. Nucleic Acids Res. 43 , W589 – W598 (2015).
CAS PubMed PubMed Central Статья Google Scholar
Алекса, А., и Раненфюрер, Дж. Анализ обогащения генетического набора с помощью topGO. Bioconductor Improv. 27 (2009 г.).
Комплексный и сравнительный фенотипический анализ коллаборативных штаммов-основателей позволяет выявить новые и известные фенотипы
Akinpelu OV, Mujica-Mota M, Daniel SJ (2014) Связан ли сахарный диабет 2 типа с нарушениями слуха? Систематический обзор и метаанализ. Ларингоскоп 124: 767–776
PubMed Google Scholar
Amrani A, Chaouloff F, Mormede P, Dardenne M, Homo-Delarche F (1994) Глюкоза, инсулин и ответы открытого поля на иммобилизацию у мышей, не страдающих ожирением, диабетом (NOD).Physiol Behav 56: 241–246
PubMed CAS Google Scholar
Benjamini Y, Hochberg Y (1995) Контроль уровня ложных открытий: практичный и мощный подход к множественному тестированию. J Roy Stat Soc 57: 289–300
Google Scholar
Bogue MA, Grubb SC, Walton DO, Philip VM, Kolishovski G, Stearns T, Dunn MH, Skelly DA, Kadakkuzha B, TeHennepe G, Kunde-Ramamoorthy G, Chesler EJ (2018) База данных феноменов мыши: интегративная база данных и набор для анализа данных эмпирического фенотипа лабораторных мышей.Нуклеиновые кислоты Res 46: D843 – D850
PubMed CAS Google Scholar
Бог М.А., Филип В.М., Уолтон Д.О., Грабб С.К., Данн М.Х., Колишовски Г., Эмерсон Дж., Мукерджи Г., Стернс Т., Хе Х., Синха В., Кадаккужа Б., Кунде-Рамамурти Г., Чеслер Э.Дж. (2019) База данных фенома мышей: репозиторий данных и набор для анализа данных о первичных фенотипах мышей. Нуклеиновые кислоты Res 48 (D1): D716 – D723
Google Scholar
Болен М., Хейс Э. Р., Болен Б., Байлоо Дж. Д., Крабб Дж. К., Уолстен Д. (2014) Влияние экспериментатора на результаты поведенческих тестов восьми инбредных линий мышей под влиянием этанола.Behav Brain Res 272: 46–54
PubMed PubMed Central CAS Google Scholar
Burgess SC, Jeffrey FM, Storey C, Milde A, Hausler N, Merritt ME, Mulder H, Holm C, Sherry AD, Malloy CR (2005) Влияние линии мышей на метаболические пути производства глюкозы после кратковременного или длительного голодание. Am J Physiol 289: E53–61
CAS Google Scholar
Кэмпбелл Дж. Х., Фостер С. М., Вишнивецкая Т., Кэмпбелл А. Г., Ян З. К., Ваймор А., Палумбо А. В., Чеслер Е. Дж., Подар М. (2012) Генетические эффекты хозяина и влияние окружающей среды на микробиоту кишечника мышей.ISME J 6: 2033–2044
PubMed PubMed Central CAS Google Scholar
Champy MF, Selloum M, Zeitler V, Caradec C, Jung B, Rousseau S, Pouilly L, Sorg T, Auwerx J (2008) Генетический фон определяет метаболические фенотипы у мышей. Геном Mamm 19: 318–331
PubMed CAS Google Scholar
Chen YG, Mathews CE, Driver JP (2018) Роль мышей NOD в исследованиях диабета 1 типа: уроки прошлого и рекомендации на будущее.Передний эндокринол 9:51
Google Scholar
Черчилль Г.А., Гатти Д.М., Мангер С.К., Свенсон К.Л. (2012) Разнообразие популяции аутбредных мышей. Геном Mamm 23: 713–718
PubMed PubMed Central Google Scholar
Courtney SM, Massett MP (2012) Идентификация QTL переносимости физической нагрузки с использованием ассоциативного картирования у инбредных мышей. Physiol Genomics 44: 948–955
PubMed PubMed Central CAS Google Scholar
Дикинсон М.Э., Фленникен А.М., Джи Х, Тебул Л., Вонг М.Д., Уайт Дж. К., Михан Т.Ф., Венингер В.Дж., Вестерберг Х., Адиссу Х., Бейкер С.Н., Бауэр Л., Браун Дж. М., Кэдл Л. Б., Кьяни Ф., Клэри D, Cleak J, Daly MJ, Denegre JM, Doe B, Dolan ME, Edie SM, Fuchs H, Gailus-Durner V, Galli A, Gambadoro A, Gallegos J, Guo S, Horner NR, Hsu CW, Johnson SJ, Kalaga С., Кейт Л.К., Ланоуэ Л., Лоусон Т.Н., Лек М., Марк М., Маршалл С., Мейсон Дж., Макэлви М.Л., Ньюбиггинг С., Наттер Л.М., Петерсон К.А., Рамирес-Солис Р., Роуленд Д.Д., Райдер Е., Самоча К.Э., Сивитт JR, Selloum M, Szoke-Kovacs Z, Tamura M, Trainor AG, Tudose I, Wakana S, Warren J, Wendling O, West DB, Wong L, Yoshiki A, International Mouse Phenotyping C, Jackson L, Infrastructure Nationale Phenomin ICdlS, Charles River L, Harwell MRC, Toronto Center for P, Wellcome Trust Sanger I, Center RB, MacArthur DG, Tocchini-Valentini GP, Gao X, Flicek P, Bradley A, Skarnes WC, Justice MJ, Parkinson HE, Moore M, Wells S, Braun RE, Svenson KL, de Angelis MH, Эро Y, Mohun T, Mallon AM, Henkelman RM, Brown SD, Adams DJ, Lloyd KC, McKerlie C, Beaudet AL, Bucan M, Murray SA (2016) Высокопроизводительное открытие новых фенотипов развития.Nature 537: 508–514
PubMed PubMed Central CAS Google Scholar
Ferris MT, Aylor DL, Bottomly D, Whitmore AC, Aicher LD, Bell TA, Bradel-Tretheway B, Bryan JT, Buus RJ, Gralinski LE, Haagmans BL, McMillan L, Miller DR, Rosenzweig E, Valdar W. , Wang J, Churchill GA, Threadgill DW, McWeeney SK, Katze MG, Pardo-Manuel de Villena F, Baric RS, Heise MT (2013) Моделирование генетической регуляции патогенеза гриппа хозяином при совместном скрещивании.PLoS Pathog 9: e1003196
PubMed PubMed Central Google Scholar
Fuchs H, Gailus-Durner V, Adler T, Pimentel JA, Becker L, Bolle I, Brielmeier M, Calzada-Wack J, Dalke C, Ehrhardt N, Fasnacht N, Ferwagner B, Frischmann U, Hans W., Holter SM, Holzlwimmer G, Horsch M, Javaheri A, Kallnik M, Kling E, Lengger C, Maier H, Mossbrugger I, Morth C, Naton B, Noth U, Pasche B, Prehn C, Przemeck G, Puk O, Racz I , Rathkolb B, Rozman J, Schable K, Schreiner R, Schrewe A, Sina C, Steinkamp R, Thiele F, Willershauser M, Zeh R, Adamski J, Busch DH, Beckers J, Behrendt H, Daniel H, Esposito I, Favor J, Graw J, Heldmaier G, Hofler H, Ivandic B, Katus H, Klingenspor M, Klopstock T., Ленгелинг A, Mempel M, Muller W, Neschen S, Ollert M, Quintanilla-Martinez L, Rosenstiel P, Schmidt J, Schreiber S, Schughart K, Schulz H, Wolf E, Wurst W, Zimmer A, Hrabe de Angelis M (2009) Немецкая клиника мышей: платформа для анализа системного фенотипа моделей мышей.Curr Pharm Biotechnol 10: 236–243
PubMed CAS Google Scholar
Fuchs H, Gailus-Durner V, Adler T, Aguilar-Pimentel JA, Becker L, Calzada-Wack J, Da Silva-Buttkus P, Neff F, Gotz A, Hans W., Holter SM, Horsch M, Kastenmuller G, Kemter E, Lengger C, Maier H, Matloka M, Moller G, Naton B, Prehn C, Puk O, Racz I, Rathkolb B, Romisch-Margl W, Rozman J, Wang-Sattler R, Schrewe A, Stoger C , Tost M, Adamski J, Aigner B, Beckers J, Behrendt H, Busch DH, Esposito I, Graw J, Illig T, Ivandic B, Klingenspor M, Klopstock T., Kremmer E, Mempel M, Neschen S, Ollert M, Schulz H, Suhre K, Wolf E, Wurst W, Zimmer A, Hrabe de Angelis M (2011) Фенотипирование мышей.Методы 53: 120–135
PubMed CAS Google Scholar
Fuchs H, Gailus-Durner V, Neschen S, Adler T, Afonso LC, Aguilar-Pimentel JA, Becker L, Bohla A, Calzada-Wack J, Cohrs C, Dewert A, Fridrich B, Garrett L, Glasl L, Gotz A, Hans W, Holter SM, Horsch M, Hurt A, Janas E, Janik D, Kahle M, Kistler M, Klein-Rodewald T, Lengger C, Ludwig T, Maier H, Marschall S, Micklich K, Moller G, Naton B, Prehn C, Puk O, Racz I, Rass M, Rathkolb B, Rozman J, Scheerer M, Schiller E, Schrewe A, Steinkamp R, Stoger C, Sun M, Szymczak W, Treise I, Vargas Panesso IL , Vernaleken AM, Willershauser M, Wolff-Muscate A, Zeh R, Adamski J, Beckers J, Bekeredjian R, Busch DH, Eickelberg O, Favor J, Graw J, Hofler H, Hoschen C, Katus H, Klingenspor M, Klopstock T , Neff F, Ollert M, Schulz H, Stoger T, Wolf E, Wurst W, Yildirim AO, Zimmer A, Hrabe de Angelis M (2012) Инновации в фенотипировании моделей мышей в Немецкой клинике мышей.Геном Mamm 23: 611–622
PubMed PubMed Central CAS Google Scholar
Gailus-Durner V, Fuchs H, Becker L, Bolle I, Brielmeier M, Calzada-Wack J, Elvert R, Ehrhardt N, Dalke C, Franz TJ, Grundner-Culemann E, Hammelbacher S, Holter SM, Holzlwimmer G, Horsch M, Javaheri A, Kalaydjiev SV, Klempt M, Kling E, Kunder S, Lengger C, Lisse T, Mijalski T., Naton B, Pedersen V, Prehn C, Przemeck G, Racz I, Reinhard C, Reitmeir P, Schneider I, Schrewe A, Steinkamp R, Zybill C, Adamski J, Beckers J, Behrendt H, Favor J, Graw J, Heldmaier G, Hofler H, Ivandic B, Katus H, Kirchhof P, Klingenspor M, Klopstock T, Ленгелинг A , Muller W, Ohl F, Ollert M, Quintanilla-Martinez L, Schmidt J, Schulz H, Wolf E, Wurst W., Zimmer A, Busch DH, de Angelis MH (2005) Знакомство с German Mouse Clinic: платформа открытого доступа для стандартизированных фенотипирование.Nat Методы 2: 403–404
PubMed CAS Google Scholar
Gailus-Durner V, Fuchs H, Adler T, Aguilar Pimentel A, Becker L, Bolle I, Calzada-Wack J, Dalke C, Ehrhardt N, Ferwagner B, Hans W, Holter SM, Holzlwimmer G, Horsch M , Джавахери А., Калльник М., Клинг Э., Ленггер С., Морт С., Моссбруггер I, Натон Б., Прен С., Пук О, Ратколб Б., Розман Дж., Шреве А., Тиле Ф, Адамски Дж., Айгнер Б., Берендт Х, Буш DH, Favor J, Graw J, Heldmaier G, Ivandic B, Katus H, Klingenspor M, Klopstock T, Kremmer E, Ollert M, Quintanilla-Martinez L, Schulz H, Wolf E, Wurst W., de Angelis MH (2009) Системный фенотипирование первой линии.Методы Mol Biol (Клифтон, Нью-Джерси) 530: 463–509
Graham JB, Thomas S, Swarts J, McMillan AA, Ferris MT, Suthar MS, Treuting PM, Ireton R, Gale M, Jr., Lund JM ( 2015) Генетическое разнообразие в совместной кросс-модели резюмирует исходы болезни, вызванной вирусом Западного Нила. mBio 6
Gralinski LE, Bankhead A, 3rd, Jeng S, Menachery VD, Proll S, Belisle SE, Matzke M, Webb-Robertson BJ, Luna ML, Shukla AK, Ferris MT, Bolles M, Chang J, Aicher L, Waters KM, Smith RD, Metz TO, Law GL, Katze MG, McWeeney S, Baric RS (2013) Механизмы тяжелого острого респираторного синдрома, вызванного коронавирусом, острого повреждения легких.mBio 4
Gralinski LE, Ferris MT, Aylor DL, Whitmore AC, Green R, Frieman MB, Deming D, Menachery VD, Miller DR, Buus RJ, Bell TA, Churchill GA, Threadgill DW, Katze MG, McMillan L , Valdar W., Heise MT, Pardo-Manuel de Villena F, Baric RS (2015) Полногеномная идентификация локусов восприимчивости к SARS-CoV с использованием совместного скрещивания. PLoS Genet 11: e1005504
PubMed PubMed Central Google Scholar
Grubb SC, Bult CJ, Bogue MA (2014) База данных феноменов мышей.Нуклеиновые кислоты Res 42: D825–834
PubMed CAS Google Scholar
Hong BN, Kang TH (2014) Различие между слуховыми электрофизиологическими ответами в моделях диабета на животных типа 1 и типа 2. Neurosci Lett 566: 309–314
PubMed CAS Google Scholar
Хорикава С., Кодама С., Танака С., Фуджихара К., Хирасава Р., Ячи Й, Шимано Х, Ямада Н., Сайто К., Соне Х (2013) Диабет и риск нарушения слуха у взрослых: метаанализ.J Clin Endocrinol Metab 98: 51–58
PubMed CAS Google Scholar
Johnson KR, Zheng QY, Noben-Trauth K (2006) Штаммовые фоновые эффекты и генетические модификаторы слуха у мышей. Brain Res 1091: 79–88
PubMed PubMed Central CAS Google Scholar
Jorgensen MS, Tornqvist KS, Hvid H (2017) Расчет дозы глюкозы для внутрибрюшинных тестов толерантности к глюкозе у худых и страдающих ожирением мышей.J Am Assoc Lab Anim Sci 56: 95–97
PubMed PubMed Central Google Scholar
Карп Н.А., Мелвин Д., Сэнгер Маус Генетикс П., Мотт Р.Ф. (2012) Надежный и чувствительный анализ фенотипов нокаута мышей. PLoS ONE 7: e52410
PubMed PubMed Central CAS Google Scholar
Карп Н.А., Михан Т.Ф., Морган Х., Мейсон Дж. К., Блейк А., Курбатова Н., Смедли Д., Якобсен Дж., Мотт Р. Ф., Айер В., Мэтьюз П., Мелвин Д. Г., Уэллс С., Фленникен А. М., Масуя Х, Вакана S, White JK, Lloyd KC, Reynolds CL, Paylor R, West DB, Svenson KL, Chesler EJ, de Angelis MH, Tocchini-Valentini GP, Sorg T, Herault Y, Parkinson H, Mallon AM, Brown SD (2015) руководящие принципы ARRIVE в базу данных in vivo.PLoS Biol 13: e1002151
PubMed PubMed Central Google Scholar
Keane TM, Goodstadt L, Danecek P, White MA, Wong K, Yalcin B, Heger A, Agam A, Slater G, Goodson M, Furlotte NA, Eskin E, Nellaker C, Whitley H, Cleak J, Janowitz D, Эрнандес-Плиего П., Эдвардс А., Белгард Т.Г., Оливер П.Л., Макинтайр Р.Э., Бхомра А., Никод Дж., Ган Х, Юань В., ван дер Вейден Л., Стюард К.А., Бала С., Сталкер Дж., Мотт Р., Дурбин Р. , Джексон И. Дж., Чехански А., Герра-Ассункао Дж. А., Донахью Л. Р., Рейнхольдт Л. Г., Пейсер Б. А., Понтинг С. П., Бирни Е., Флинт Дж., Адамс Д. Д. (2011) Геномная вариация мыши и ее влияние на фенотипы и регуляцию генов.Nature 477: 289–294
PubMed PubMed Central CAS Google Scholar
Kelada SN (2016) QTL плетизмографического фенотипа у мышей до и после аллергической сенсибилизации и заражения. G3 6: 2857–2865
PubMed CAS Google Scholar
Kelada SN, Aylor DL, Peck BC, Ryan JF, Tavarez U, Buus RJ, Miller DR, Chesler EJ, Threadgill DW, Churchill GA, Pardo-Manuel de Villena F, Collins FS (2012) Генетический анализ гематологических параметры в зарождающихся линиях коллаборативного креста.Genes Genomes Genetics 2: 157–165
PubMed Google Scholar
Kleinert M, Clemmensen C, Hofmann SM, Moore MC, Renner S, Woods SC, Huypens P, Beckers J, de Angelis MH, Schurmann A, Bakhti M, Klingenspor M, Heiman M, Cherrington AD, Ristow M, Lickert H, Wolf E, Havel PJ, Muller TD, Tschop MH (2018) Животные модели ожирения и сахарного диабета. Nat Rev Endocrinol 14: 140–162
PubMed Google Scholar
Кулесская Н., Войкар В. (2014) Оценка тревожного поведения мышей в светло-темном боксе и арене открытого поля: роль оборудования и процедуры.Physiol Behav 133: 30–38
PubMed CAS Google Scholar
Курбатова Н., Мейсон Дж. К., Морган Х., Михан Т.Ф., Карп Н.А. (2015) PhenStat: набор инструментов для стандартизованного анализа высокопроизводительных фенотипических данных. PLoS ONE 10: e0131274
PubMed PubMed Central Google Scholar
Lad HV, Liu L, Paya-Cano JL, Parsons MJ, Kember R, Fernandes C, Schalkwyk LC (2010) Тестирование поведенческой батареи: оценка и результаты поведения у 8 инбредных линий мышей.Physiol Behav 99: 301–316
PubMed CAS Google Scholar
Leist SR, Pilzner C, van den Brand JM, Dengler L, Geffers R, Kuiken T., Balling R, Kollmus H, Schughart K (2016) Инфекция гриппом h4N2 совместных перекрестных штаммов-основателей выявляет сильно расходящиеся ответы хозяина и определяет уникальный фенотип у мышей CAST / EiJ. BMC Genomics 17: 143
PubMed PubMed Central Google Scholar
Lilue J, Doran AG, Fiddes IT, Abrudan M, Armstrong J, Bennett R, Chow W, Collins J, Collins S, Chechanski A, Danecek P, Diekhans M, Dolle DD, Dunn M, Durbin R, Earl Д., Фергюсон-Смит А., Фличек П., Флинт Дж., Фрэнкиш А., Фу Б., Герштейн М., Гилберт Дж., Гудштадт Л., Харроу Дж., Хоу К., Ибарра-Сориа Х, Колмогоров М., Леллиотт С.Дж., Логан Д.В., Лавленд Дж. , Mathews CE, Mott R, Muir P, Nachtweide S, Navarro FCP, Odom DT, Park N, Pelan S, Pham SK, Quail M, Reinholdt L, Romoth L, Shirley L, Sisu C, Sjoberg-Herrera M, Stanke M , Steward C, Thomas M, Threadgold G, Thybert D, Torrance J, Wong K, Wood J, Yalcin B, Yang F, Adams DJ, Paten B, Keane TM (2018) Шестнадцать различных эталонных геномов лабораторных мышей определяют штамм-специфические гаплотипы и новые функциональные локусы.Nat Genet 50: 1574–1583
PubMed PubMed Central CAS Google Scholar
Logan RW, Robledo RF, Recla JM, Philip VM, Bubier JA, Jay JJ, Harwood C, Wilcox T, Gatti DM, Bult CJ, Churchill GA, Chesler EJ (2013) Высокоточное генетическое картирование поведенческих черт в разнообразии популяции беспородных мышей. Гены поведения мозга 12: 424–437
PubMed PubMed Central CAS Google Scholar
Mandillo S, Tucci V, Holter SM, Meziane H, Banchaabouchi MA, Kallnik M, Lad HV, Nolan PM, Ouagazzal AM, Coghill EL, Gale K, Golini E, Jacquot S, Krezel W, Parker A, Riet F, Schneider I, Marazziti D, Auwerx J, Brown SD, Chambon P, Rosenthal N, Tocchini-Valentini G, Wurst W. (2008) Надежность, устойчивость и воспроизводимость в поведенческом фенотипировании мышей: межлабораторное исследование.Physiol Genomics 34: 243–255
PubMed PubMed Central Google Scholar
О’Коннор А., Кизон П.М., Олбрайт Дж. Э., Лин Ф. Т., Беннетт Б. Дж. (2014) Реакция кардиометаболической микробиоты на диету зависит от генетики хозяина. Геном Mamm 25: 583–599
PubMed PubMed Central CAS Google Scholar
Oellrich A, Meehan TF, Parkinson H, Sarntivijai S, White JK, Karp NA (2016) Сообщение о фенотипах в моделях мышей при рассмотрении размера тела как потенциального фактора, мешающего.Дж Биомед Семант 7: 2
Google Scholar
Pan HJ, Agate DS, King BL, Wu MK, Roderick SL, Leiter EH, Cohen DE (2006) Полиморфизм в новозеландских инбредных линиях мышей, который инактивирует белок-переносчик фосфатидилхолина. FEBS Lett 580: 5953–5958
PubMed PubMed Central CAS Google Scholar
Panganiban CH, Barth JL, Darbelli L, Xing Y, Zhang J, Li H, Noble KV, Liu T, Brown LN, Schulte BA, Richard S, Lang H (2018) Нарушение регуляции сотрясения связывания РНК, вызванное шумом белки способствуют демиелинизации слухового нерва и потере слуха.J Neurosci 38: 2551–2568
PubMed PubMed Central CAS Google Scholar
Персиваль С.Дж., Либертон Д.К., Пардо-Мануэль де Вильена Ф., Спритц Р., Маркуцио Р., Халльгримссон Б. (2016) Генетика черепно-лицевой морфологии мышей: диаллельный анализ восьми основателей Коллаборативного креста. J Anat 228: 96–112
PubMed Google Scholar
Phillippi J, Xie Y, Miller DR, Bell TA, Zhang Z, Lenarcic AB, Aylor DL, Krovi SH, Threadgill DW, de Villena FP, Wang W, Valdar W, Frelinger JA (2014) Использование новых совместных крестик, чтобы проверить иммунную систему.Genes Immun 15: 38–46
PubMed CAS Google Scholar
Рид Д.Р., Лоулер М.П., Тордофф М.Г. (2008) Сниженная масса тела — частый эффект нокаута гена у мышей. BMC Genet 9: 4
PubMed PubMed Central Google Scholar
Робертс А., Пардо-Мануэль де Вильена Ф, Ван В., Макмиллан Л., Тредгилл Д. В. (2007) Архитектура полиморфизма генетических ресурсов мышей, выясненная с использованием данных повторного секвенирования генома: последствия для открытия QTL и системной генетики.Геном Mamm 18: 473–481
PubMed PubMed Central CAS Google Scholar
Rogers DC, Peters J, Martin JE, Ball S, Nicholson SJ, Witherden AS, Hafezparast M, Latcham J, Robinson TL, Quilter CA, Fisher EM (2001) SHIRPA, протокол для поведенческой оценки: проверка для продольной исследование неврологической дисфункции на мышах. Neurosci Lett 306: 89–92
PubMed CAS Google Scholar
Rutledge H, Aylor DL, Carpenter DE, Peck BC, Chines P, Ostrowski LE, Chesler EJ, Churchill GA, de Villena FP, Kelada SN (2014) Генетическая регуляция Zfp30, CXCL1 и нейтрофильного воспаления в легких мыши .Генетика 198: 735–745
PubMed PubMed Central CAS Google Scholar
Saul MC, Philip VM, Reinholdt LG, Chesler EJ (2019) Популяции мышей с большим разнообразием по сложным признакам. Тенденции Genet 35: 501–514
PubMed CAS Google Scholar
Сковилль Д.К., Уайт СС, Ботта Д., МакКонначи Л.А., Задворни М.Э., Шмук СК, Ху Х, Гао Х, Ю Дж., Диллс Р.Л., Шеппард Л., Делани М.А., Гриффит В.С., Бейер Р.П., Зангар Р.С., Фунты JG, Faustman EM, Kavanagh TJ (2015) Восприимчивость к воспалению легких, индуцированному квантовыми точками, широко различается среди совместных линий мышей-кросс-основателей.Toxicol Appl Pharmacol 289: 240–250
PubMed PubMed Central CAS Google Scholar
Smith CM, Proulx MK, Olive AJ, Laddy D, Mishra BB, Moss C, Gutierrez NM, Bellerose MM, Barreira-Silva P, Phuah JY, Baker RE, Behar SM, Kornfeld H, Evans TG, Beamer G , Sassetti CM (2016) Восприимчивость к туберкулезу и защита от вакцины независимо контролируются генотипом хозяина. mBio 7
Svenson KL, Von Smith R, Magnani PA, Suetin HR, Paigen B, Naggert JK, Li R, Churchill GA, Peters LL (2007) Измерения множественных признаков у 43 инбредных линий мышей отражают фенотипическое разнообразие, характерное для человеческие популяции.J Appl Physiol (1985) 102: 2369–2378
CAS Google Scholar
Svenson KL, Gatti DM, Valdar W., Welsh CE, Cheng R, Chesler EJ, Palmer AA, McMillan L, Churchill GA (2012) Генетическое картирование с высоким разрешением с использованием аутбредной популяции мышей. Генетика 190: 437–447
PubMed PubMed Central CAS Google Scholar
Takahashi A, Kato K, Makino J, Shiroishi T, Koide T (2006) Многомерный анализ временных описаний поведения в открытом поле у штаммов мышей дикого происхождения.Behav Genet 36: 763–774
PubMed Google Scholar
The Collaborative Cross Consortium (2012) Архитектура генома совместной генетической эталонной популяции мышей. Генетика 190: 389–401
Google Scholar
Веред К., Дюррант С., Мотт Р., Иракский Ф.А. (2014) Восприимчивость к инфекции klebsiella pneumonaie у мышей совместного скрещивания — сложный признак, контролируемый по крайней мере тремя локусами, действующими в разные моменты времени.BMC Genomics 15: 865
PubMed PubMed Central Google Scholar
West BT, Welch KB, Galecki A (2014) Линейные смешанные модели: практическое руководство с использованием статистического программного обеспечения, 2-е изд. Чепмен и Холл / CRC, Нью-Йорк
Google Scholar
Вонг А.А., Браун Р.Э. (2006) Обнаружение зрения, распознавание образов и острота зрения у 14 линий мышей. Genes Brain Behav 5: 389–403
PubMed CAS Google Scholar
Zhang J, Malo D, Mott R, Panthier JJ, Montagutelli X, Jaubert J (2018) Идентификация новых локусов, участвующих в восприимчивости хозяина к Salmonella Typhimurium у мышей совместного скрещивания.BMC Genomics 19: 303
PubMed PubMed Central Google Scholar
Zheng QY, Johnson KR, Erway LC (1999) Оценка слуха у 80 инбредных линий мышей с помощью порогового анализа ABR. Послушайте Res 130: 94–107
PubMed PubMed Central CAS Google Scholar
Границы | Автоматизированные методы позволяют проводить прямые вычисления фенотипических описаний для предсказания нового гена-кандидата
Предпосылки
Фенотипы включают в себя огромное количество важной и полезной информации о растениях, потенциально включая состояния, связанные с приспособленностью, болезнями и сельскохозяйственной ценностью.Они представляют собой материал, на который воздействует естественный и искусственный отбор для повышения приспособленности или достижения желаемых качеств, соответственно. Определение того, какие гены связаны с интересующими чертами, и понимание природы этих отношений имеет решающее значение для манипулирования фенотипами. Когда выявлены причинные аллели интересующих фенотипов, они могут быть отобраны в популяциях, нацелены на делецию или использованы в качестве трансгенов для введения желаемых признаков внутри и между видами.Процесс идентификации генов-кандидатов и конкретных аллелей, связанных с интересующим признаком, называется предсказанием генов-кандидатов.
Гены со сходными последовательностями часто имеют общие биологические функции и поэтому могут создавать похожие фенотипы. Это одна из причин, по которой алгоритмы поиска сходства последовательностей, такие как BLAST (Altschul et al., 1990), так полезны для предсказания генов-кандидатов. Однако сходные фенотипы также можно отнести к функции генов, у которых нет сходства последовательностей.Вот как гены, кодирующие белок, которые участвуют в разных этапах одного и того же метаболического пути, или факторы транскрипции, участвующие в регуляции экспрессии генов, вносят свой вклад в общие фенотипы. Например, отключение любого из многих генов, участвующих в пути антоцианов кукурузы, может привести к изменениям пигмента (обзор Sharma et al., 2011). Эта концепция смоделирована на рисунке 1, где, в частности, поиск на основе последовательностей с геном 1 в качестве запроса может возвращать только гены с аналогичными последовательностями, но запрос фенотипов, сходных с фенотипами, связанными с геном 1, возвращает множество дополнительных генов-кандидатов.
Рисунок 1 Концептуальное сравнение запросов с последовательностью гена или связанным с ней фенотипическим описанием. Гены показаны в виде белых овалов . Способы поиска родственных генов показаны как светло-серыми прямоугольниками . Серые пунктирные стрелки указывают путь от запрашиваемого гена к набору генов, которые возвращаются в результате поиска. Сплошные черные стрелки указывают отношения между генами в биологическом пути или в сети регуляции генов. Пунктирные черные стрелки указывают отношения между путем или сетью и результирующим фенотипом.
Высокопроизводительные и вычислительные методы фенотипирования в основном основаны на сенсорах и изображениях (Fahlgren et al., 2015). Эти методы позволяют создавать стандартизированные наборы данных, например, анализировать изображение, извлекать данные и запрашивать эти данные (Green et al., 2012; Gehan et al., 2017; Miller et al., 2017). ). Однако, хотя такие методы позволяют сравнивать фенотипическую информацию между физически похожими растениями, их способность передавать эти знания между физически разнородными видами ограничена.Например, такие характеристики, как угол наклона листьев, сильно различаются у разных видов, и поэтому их нельзя сравнивать напрямую. Более того, там, где общие пути и процессы сохраняются на больших эволюционных дистанциях, может быть трудно идентифицировать эквивалентные фенотипы. McGary et al. (2010) называют эти неочевидные общие фенотипы фенологами. Между видами фенологи могут проявляться как эквивалентные свойства в разрозненных биологических структурах (Braun et al., 2018). Например, мутанты KIN-13A Arabidopsis и мутанты KIF2A мыши демонстрируют повышенное ветвление в одноклеточных структурах, но по отношению к нейронам мыши (Homma et al., 2003) и по трихомам у Arabidopsis (Lu et al., 2004). Взятые вместе, способность вычислять фенотипические описания для идентификации фенологов внутри и между видами может помочь в идентификации новых генов-кандидатов, которые не могут быть идентифицированы только методами на основе последовательностей и которые не могут быть идентифицированы с помощью анализа изображений .
Для идентификации фенологов некоторые методы основываются на поиске общих ортологов между наборами причинных генов (McGary et al., 2010; Woods et al., 2013). Например, McGary et al. (2010) идентифицировали фенологическую связь между «аномальным развитием сердца» у мышей и «дефектным ответом на красный свет» у Arabidopsis , идентифицировав четыре ортологичных гена между наборами известных причинных генов у каждого вида. Однако эти методы неприменимы, когда известный причинный набор генов для одного или другого фенотипа невелик или отсутствует. В этих случаях использование описаний на естественном языке для идентификации фенологов позволяет избежать этой проблемы, полагаясь только на характеристики фенотипов как таковых.Эти фенотипические описания являются богатым источником информации, которая, если ее использовать для идентификации пар фенологов, может позволить идентифицировать новые гены-кандидаты, потенциально участвующие в генерации фенотипов, выходящих за рамки того, что уже было описано.
К сожалению, вычислить описания фенотипов непросто. Текстовые описания фенотипов, представленные в литературе и в онлайн-базах данных, нерегулярны, потому что представления на естественном языке даже очень похожих фенотипов могут сильно варьироваться.Это делает надежную количественную оценку сходства фенотипов особенно сложной задачей (Thessen et al., 2012; Braun et al., 2018). Чтобы представить фенотипы вычислимым образом, исследователи недавно начали переводить и стандартизировать описания фенотипов в утверждения «сущность-качество» (EQ), состоящие из терминов онтологии, где сущность (например, «лист») модифицируется с помощью качества (например, « увеличенная длина »; Mungall et al., 2010). Используя этот формализм, сложные фенотипы представлены множеством операторов EQ.Например, несколько операторов EQ требуются для представления карликовости, где среди прочего могут использоваться пары сущность и качество («высота растения», «уменьшенный») и («ширина листа» «увеличенный»). Каждый из этих фенотипических компонентов более общего фенотипа называется «фен». Поскольку и сущности, и качества представлены терминами из биологических онтологий (фиксированные словари, организованные в виде иерархических понятий в направленном ациклическом графе), количественная оценка сходства между двумя фенотипами, которые были переведены в выражения EQ, может быть выполнена с использованием показателей сходства на основе графа (Hoehndorf и другие., 2011; Слимани, 2013). Такие методы оценки семантического сходства, основанные на иерархическом расположении понятий в графе, уже давно используются в области обработки естественного языка (NLP; например, Resnik, 1999) и, применительно к биологическим онтологиям, были полезны в приложениях из кластеризации генов. функциональные аннотации для визуализации данных (Supek et al., 2011) для оценки функционального сходства между ортологичными генами (Altenhoff et al., 2016).
Oellrich, Walls et al.(2015) разработали Plant PhenomeNET, ресурс на основе EQ, в основном состоящий из сети сходства фенотипов, содержащей фенотипы шести различных модельных видов растений, а именно Arabidopsis ( Arabidopsis thaliana ), кукурузы ( Zea mays ssp. mays ), томат ( Solanum lycopersicum ), рис ( Oryza sativa ), Medicago ( Medicago truncatula ) и соя ( Glycine max ). Их анализ показал, что метод, разработанный Hoehndorf et al.(2011) можно использовать для восстановления известных ассоциаций генотипа и фенотипа для растений. Авторы обнаружили, что очень похожие фенотипы в сети (фенологи), вероятно, имеют общие причинные гены, которые являются ортологами или участвуют в одних и тех же биологических путях. При построении сети текстовые утверждения, содержащие каждый фенотип, вручную преобразовывались в утверждения EQ, в основном состоящие из терминов из онтологии фенотипа и признака (PATO; Gkoutos et al., 2005), онтологии растений (PO; Cooper et al., 2013), Онтология генов (GO; Ashburner et al., 2000) и Химические объекты, представляющие биологический интерес (ChEBI; Hastings et al., 2013).
Успех этого пилотного проекта по фенотипу растений был обнадеживающим, но масштабирование до вычисления всех доступных фенотипических данных для каждого из шести видов не было разумной целью, учитывая, что сбор данных для этого пилотного проекта занял приблизительно 2 года и охватил только фенотипы доминантных аллелей для 2747 генов шести видов. В частности, человеческий перевод текстовых заявлений в выражения EQ является наиболее трудоемким аспектом создания сетей сходства фенотипов с использованием этого метода.Автоматизация этого перевода обещает увеличить скорость создания и расширения таких сетей. Известные усилия по автоматизации этого процесса включают Semantic Charaparser (Cui, 2012; Cui et al., 2015), который извлекает символы (сущности) и их соответствующие состояния (качества) после этапа курирования, который включает присвоение терминов категориям и последующее отображение этих символов. и состояния к операторам EQ, построенным на основе входных онтологий. Другие существующие инструменты аннотации, такие как NCBO Annotator (Musen et al., 2012) и NOBLE Coder (Цейтлин и др., 2016) полностью автоматизированы, полагаясь только на онтологии ввода. Оба отображают слова во входном тексте в термины онтологии без навязывания структуры оператора EQ. Также были разработаны современные подходы машинного обучения к аннотированию текста с помощью онтологических терминов (Hailu et al., 2019). Их можно обучить с помощью набора данных, такого как Colorado Richly Annotated Full-Text corpus (CRAFT; Bada et al., 2012), но их нелегко перенести на онтологии, которые не представлены в обучающем наборе.
Помимо использования методов, основанных на онтологии, сходство между текстовыми описаниями фенотипов также можно количественно оценить с помощью методов НЛП, таких как обработка каждого описания как набора слов и сравнение наличия или отсутствия этих слов между описаниями или использование Инструменты на основе нейронных сетей, такие как Doc2Vec, для встраивания описаний в абстрактные многомерные числовые векторы, между которыми затем могут быть легко применены метрики сходства (Mikolov et al., 2013; Le and Mikolov, 2014).Концептуально этот процесс включает преобразование описаний на естественном языке в местоположения в пространстве, так что описания, которые находятся рядом друг с другом, интерпретируются как имеющие большое сходство, а те, которые находятся на расстоянии, имеют низкое сходство.
В этой работе мы демонстрируем, что автоматизированные методы генерации вычислимых представлений естественного языка могут быть применены к набору данных фенотипических описаний для создания биологически значимых сетей сходства фенотипов.См. Рисунок 2 для обзора того, как сети сходства фенотипов генерируются с помощью вычислений в качестве выходных данных, когда текстовые описания предоставляются в качестве входных данных. Сначала мы покажем, что эти вычислительные методы ограничены в своей способности точно воспроизводить аннотации и соответствующие сети сходства фенотипов, созданные с помощью ручного курирования. Однако впоследствии мы покажем, что созданная вручную сеть не превосходит сети, построенные с использованием чисто вычислительных подходов, в задачах биологической значимости в масштабе всего набора данных, таких как организация генов по функциям и прогнозирование принадлежности к биохимическим путям.Что наиболее важно, мы обсуждаем, как теперь мы можем использовать эти вычислительные подходы для автоматического создания новых наборов данных, необходимых для выявления фенотипических сходств и прогнозирования функции генов внутри и между видами, не требуя трудоемкого и дорогостоящего ручного курирования.
Рисунок 2 Обзор вычислительных конвейеров, используемых здесь для создания сетей сходства фенотипов из текстовых описаний фенотипов. Закругленные белые прямоугольники представляют данные в виде текстовых описаний в качестве входных или сетевых узлов в качестве выходных. Закругленные черные прямоугольники представляют промежуточные формы данных, которые являются вычислимыми представлениями текстовых описаний. Это позволяет применять количественные показатели сходства. Серые прямоугольники представляют вычислительные методы, выполняемые на каждом этапе. Однонаправленные стрелки представляют поток данных через каждый конвейер. Двуглавые стрелки представляют ребра между узлами в результирующих сетях подобия. Значения рядом с двуглавыми стрелками указывают на величину фенотипического сходства.Для каждого вычислимого представления создается одна выходная сеть, но здесь показан только один пример.
Методы
Набор данных фенотипических описаний и проверенных утверждений EQ
Сеть попарного фенотипического сходства, описанная Oellrich, Walls et al. (2015) был построен на основе набора данных описаний фенотипов шести различных модельных видов растений ( A. thaliana , Z. mays ssp. mays , S. lycopersicum , O.sativa , M. truncatula и G. max ). В этой работе описание каждого фенотипа было разделено на одно или несколько отдельных утверждений, описывающих индивидуальные фенены, каждый из которых сопоставлен ровно с одним тщательно подобранным утверждением EQ (Таблица 1). Операторы EQ в этом наборе данных были в основном построены на основе терминов, присутствующих в PATO, PO, GO и ChEBI. Для этой работы мы использовали этот существующий набор данных в качестве источника генов и связанных фенотипических описаний, на которых можно было протестировать автоматизированные методы оценки сетей сходства между фенотипами и использовать полученные сети сходства фенотипов для проведения сравнительного анализа всего набора данных для прогнозирования функции генов.
Таблица 1 Описание Oellrich, Walls et al. (2015) с точки зрения количества описаний фенотипов, описаний фенотипов и утверждений EQ.
Вычислительная генерация выражений EQ из фенотипических описаний
Для каждого фенотипа и описания фенотипа в наборе данных мы с помощью вычислений сгенерировали соответствующие выражения EQ без вмешательства человека. Для этого термины сначала были аннотированы к каждому текстовому описанию, а затем объединены для формирования полных операторов EQ.Для сопоставления терминов онтологии с текстовыми описаниями использовались два различных существующих вычислительных инструмента и простой метод машинного обучения. В частности, это были NCBO Annotator и NOBLE Coder, которые представляют собой инструменты для сопоставления терминов онтологии с конкретными словами в тексте, и наивный байесовский классификатор слов, который присваивает термины описаниям на основе наблюдаемой частоты совпадения терминов и слов. появление в наборе обучающих данных. Oellrich, Walls et al. (2015) набор данных с описаниями и тщательно подобранными утверждениями EQ был разделен на четыре группы, так что любые три группы набора данных использовались для обучения модели Наивного Байеса, которая затем применялась к оставшейся группе.Результатом применения этих трех методов аннотации стал набор терминов онтологии из PATO, PO, GO и ChEBI, назначенных каждому текстовому описанию. Затем термины были объединены для формирования полных операторов EQ путем присвоения корневых терминов по умолчанию, где ни один из них не был сопоставлен, например, термин объекта целое предприятие (PO: 0000003), и организации сопоставленных терминов по различным ролям оператора EQ путем удаления перекрывающихся термины и автоматическое применение композиционных правил, используемых кураторами в Oellrich, Walls et al.(2015). Например, эти правила включают тот факт, что термины ChEBI не могут быть первичным объектом. Утверждения EQ были оценены на основе того, насколько хорошо термины согласованы с текстовым описанием, к которому они были аннотированы, так что наиболее подходящие утверждения EQ для каждого текстового описания были выведены и использованы ниже по потоку для создания сетей сходства фенотипов. См. Раздел «Дополнительные методы» для более подробного описания этого процесса.
Вычислительная генерация числовых векторов из фенотипических описаний
Помимо генерации операторов EQ для каждого фенотипа и описания фенотипа в наборе данных, Doc2Vec использовался для генерации числовых векторов для каждого описания.Использовалась модель, предварительно обученная в Википедии (Lau and Baldwin, 2016). В этих встраиваемых документах позиции в векторе относятся не к наличию конкретных слов, а к абстрактным функциям, усвоенным моделью. Для каждого векторного представления использовался размер 300, который является фиксированным размером вектора предварительно обученной модели. Кроме того, для каждого описания были сгенерированы векторы с использованием наборов слов и представлений текста. Для этих методов каждая позиция в векторе относится к определенному слову в словаре.Каждый элемент вектора с набором слов относится к количеству этого слова в описании, а каждый элемент вектора с набором слов является двоичным значением, указывающим наличие или отсутствие слова. В случаях, когда описания фенотипа использовались вместо описаний фенотипов, описания были объединены перед внедрением для получения единого вектора.
Создание сетей генов и фенотипов
Oellrich, Walls et al. (2015) разработали сеть с фенотипами в качестве узлов и сходством между ними в качестве границ для всех фенотипов в наборе данных.Для каждого типа текстовых представлений, которые мы создали с помощью вычислительных методов, были построены сопоставимые сети. Для представлений операторов EQ подобие Жаккара либо учитывает структуру и порядок терминов в операторе EQ (называемое метрикой S 1 ), либо игнорирует структуру и обрабатывает термины онтологии в операторе EQ как неупорядоченный набор. (называемая метрикой S 2 ) использовались для определения значений границ. См. Раздел «Дополнительные методы» для более подробного описания этих показателей сходства.Для векторных представлений, созданных с использованием Doc2Vec и набора слов, использовалось косинусное сходство. Для векторных представлений, созданных с использованием набора слов, использовалось подобие Жаккара. Эти сети считаются одновременно сетями сходства генов и фенотипов, потому что каждый фенотип в наборе данных соответствует определенному причинному гену, а узел в сети представляет как этот причинный ген, так и родственный ему фенотип. Однако два описания фенотипа, соответствующие одному и тому же гену, сохраняются как два отдельных узла в сети, поэтому, хотя каждый узел представляет уникальную пару ген / фенотип, один ген может быть представлен в более чем одном узле.
Результаты
Производительность вычислительных методов при воспроизведении вручную отобранных аннотаций
Мы проверили способность методов вычислительной семантической аннотации назначать термины онтологии, аналогичные тем, которые выбирают кураторы для описаний фенотипов и фенотипов в Oellrich, Walls et al. (2015) набор данных. В частности, термины онтологии, отображаемые каждым методом в конкретное описание, сравнивались с терминами, присутствующими в операторе (ах) EQ, которые были созданы вручную для того же описания.Для оценки эффективности использовались показатели частичной точности (PP) и частичного отзыва (PR), а также среднее гармоническое этих значений (PF 1 ) в качестве сводной статистики (таблица 2). Метрики PP и PR применялись, как в Dahdul et al. (2018); см. раздел «Дополнительные методы» для подробного описания этих показателей.
Таблица 2 Показатели производительности для методов семантической аннотации.
NOBLE Coder и NCBO Annotator обычно производили семантические аннотации, более похожие на вручную подобранный набор данных, с использованием описаний фенотипов в качестве входных данных, чем с использованием набора описаний фенотипов в качестве входных данных, что соответствует всем онтологиям.Мы сочли это противоречащим интуиции, потому что описания фенов более напрямую связаны с отдельными операторами EQ с точки зрения семантического содержания. Однако набор терминов целевой онтологии, считающихся правильными, больше в случае описаний фенотипов, потому что этот набор терминов включает в себя все термины в любых утверждениях EQ, производных от этого фенотипа, а не один оператор EQ, который может способствовать этому измеряемому увеличению как частичный отзыв, так и частичная точность. Учет синонимов и связанных слов, сгенерированных с помощью моделей Word2Vec, увеличил PR в случае конкретных методов аннотации, поскольку порог схожести слов был снижен (с 1.От 0 до 0,5), но не увеличивал PF1 ни в каком случае из-за соответствующих потерь в PP (дополнительный рисунок 1).
NOBLE Coder и NCBO Annotator работали сравнимо в случае каждого типа текстового описания и онтологии, при этом NOBLE Coder использовал параметр точного соответствия, немного превосходя другой метод аннотации в отношении этих конкретных показателей для этих конкретных описаний. Оба они превзошли наивный байесовский классификатор, для которого производительность значительно упала для онтологий с меньшим относительным представлением в наборе данных (GO и ChEBI), как и следовало ожидать.Когда результаты были агрегированы, увеличение частичного отзыва для терминов PATO, PO и GO по сравнению с максимальным отзывом, достигнутым любым отдельным методом, указывает на то, что тщательно отобранные термины, которые были вызваны каждым методом, не полностью перекрывались. Это ожидаемо, учитывая, что различные методы, используемые для семантической аннотации, вызывали целевые (курируемые) термины онтологии в разной степени, что измеряется сходством Жаккара заданного целевого термина с ближайшим предсказанным термином, аннотированным этим конкретным методом.Эти наборы полученных сходств с целевыми терминами были сопоставимы между NCBO Annotator и NOBLE Coder ( ρ = 0,84 с описанием фенотипа и ρ = 0,86 с описанием фенотипа) и различались между этими методами и классификатором Наивного Байеса ( ρ <0,10 в обоих случаях для любого типа описания) с использованием ранговой корреляции Спирмена с поправкой на связи.
Эти результаты показывают, что автоматизированные методы аннотации (NCBO Annotator, NOBLE Coder и Naïve Bayes classifier) не воспроизводят точно такие же аннотации терминов онтологии, выбранные вручную для каждого фенотипического описания, как ожидалось.Учитывая этот результат, мы затем оценили, как эти различия между вручную созданными аннотациями и компьютерно сгенерированными аннотациями трансформируются в различия между сетями фенотипического сходства на основе этих аннотаций.
Сравнение вычислительных сетей с вручную созданной сетью
Oellrich, Walls et al. (2015) разработали сеть с парами фенотип / ген в качестве узлов и сходством между ними в качестве границ для всех фенотипов в наборе данных. В этой работе сопоставимые сети были построены для одного и того же набора данных с использованием ряда вычислительных подходов для представления фенотипов и описаний фенотипов, а также для прогнозирования сходства.Для целей этой оценки сеть построена на основе вручную подобранных утверждений EQ и описана в Oellrich, Walls et al. (2015) считается золотым стандартом, с которым сравнивается каждая созданная нами сеть. Вычислительные сети и сети золотого стандарта сравнивались с использованием метрики F 1 для оценки сходства в предсказанных парах фенологов в диапазоне k значений, где k — допустимое количество пар фенологов, предсказанное сетями ( k наиболее ценных ребра).Результаты представлены через k = 583 971, что представляет собой количество отличных от нуля сходств между фенотипами в сети золотого стандарта, и были повторены с использованием описаний фенотипов и фенотипов в качестве входных данных для вычислительных методов (рис. 3). Простейшие методы НЛП для оценки сходства (набор слов и набор слов) последовательно повторяли сеть золотого стандарта с использованием описаний фенотипов, тогда как метод встраивания документов с использованием Doc2Vec превзошел эти методы для значений k ≤ 200000 на основе описаний фен.Различия в производительности каждого метода устойчивы до 80% подвыборки фенотипов, присутствующих в наборе данных.
Рис. 3 Сравнение пар фенологов, идентифицированных с помощью методов прогнозирования, по сравнению с данными Oellrich, Walls et al. (2015) набор данных. Ось x указывает количество пар фенологов (ребра с наивысшим значением в сети фенотипического сходства) в каждой точке. Стандартное отклонение повторной выборки с 80% фенотипов в наборе данных (сетевые узлы) обозначены лентами для каждого метода.Описания фен ( слева, ) или описания фенотипа ( справа, ) использовались в качестве ввода текста для каждого конкретного метода.
Эти результаты показывают, что вычислительные методы не точно воспроизводят сеть фенотипического сходства, построенную на основе вручную подобранных операторов EQ. Однако это не обязательно означает, что созданная вручную сеть по своей сути более значима с биологической точки зрения. Чтобы оценить, насколько полезна каждая сеть в биологическом контексте, мы затем сравнили, как вручную созданная сеть и каждая вычислительная сеть выполняли задачу сортировки генов по функциональным группам.
Вычислительные методы превосходят ручное курирование функциональной категоризации генов в
ArabidopsisLloyd and Meinke (2012) ранее организовали набор из генов Arabidopsis с сопутствующими описаниями фенотипов в функциональную иерархию групп (например, «морфологические») , классы (например, «репродуктивный») и, наконец, подмножества (например, «цветочные») в порядке от наиболее общего к наиболее частному. См. Дополнительную таблицу 1 в Lloyd and Meinke (2012) для полной спецификации этой иерархии, к которой были отнесены гены, и дополнительную таблицу 2 в Lloyd and Meinke (2012) для сопоставления генов и этого иерархического словаря.Oellrich, Walls et al. (2015) позже использовали этот набор генов и фенотипов для проверки качества своего набора данных вручную подобранных утверждений EQ, сообщая о среднем сходстве фенотипов (переведенных в утверждения EQ), которые принадлежали к одному и тому же функциональному подмножеству. Мы использовали ту же категоризацию функциональной иерархии и аналогичный подход для оценки полезности компьютерно сгенерированных представлений фенотипов для правильной категоризации функций соответствующих генов и для сравнения этой полезности с набором данных вручную подобранных операторов EQ.Для каждого класса и подмножества в иерархии среднее сходство между любыми двумя фенотипами, связанными с генами в этом классе или подмножестве (среднее «в пределах»), было количественно оценено с использованием каждого вычислимого представления интереса и сравнено со средним сходством между фенотипом, связанным с ген в этом классе или подмножестве и один вне его (среднее «между»), количественно выраженный в терминах стандартного отклонения распределения всех оценок сходства, полученных для каждого данного метода. Разница между средним значением «внутри» и средним значением «между» (называемым здесь индексом согласованности) для каждой функциональной категории для каждого метода указывает на способность этого метода генерировать сильный сигнал сходства для фенотипов в этом наборе данных, которые разделяют эту функцию ( Рисунок 4).В случае этих данных большинство вычислительных методов, использующих описания фенотипа или фенотипа в качестве входного текста, смогли воспроизвести сигнал, присутствующий в сети. Oellrich, Walls et al. (2015) сгенерированы из вручную подобранных операторов EQ, а простейшие методы НЛП (набор слов и набор слов) дали наиболее последовательный сигнал.
Рисунок 4 Тепловая карта индекса согласованности. Показана разница между средним сходством для двух фенотипов внутри подмножества и одного фенотипа внутри и одного вне его для каждого функционального подмножества, определенного в наборе данных из фенотипов Arabidopsis , и для каждого метода количественной оценки сходства между фенотипами с более темными клетками указывает на более высокую согласованность в подмножестве.Различия измеряются в стандартных отклонениях распределений сходства, полученных для каждого метода. Значение сокращений подмножества указано в дополнительной таблице 1 Lloyd and Meinke (2012). Методы перечислены на слева . Входной текст для расчета сходства между фенотипами был получен либо из описаний фенотипов ( верхний, ), либо из описаний фенов ( нижний ). Крайний правый столбец на тепловой карте относится к среднему индексу согласованности для данного метода по всем подмножествам.
Для более непосредственного сравнения каждого метода в общей задаче классификации сети, построенные из тщательно подобранных операторов EQ, и сети, созданные с использованием каждого вычислительного метода, использовались для итеративной классификации каждого фенотипа Arabidopsis на классы и подмножества. Это было достигнуто путем удаления одного фенотипа за раз и исключения оставшихся фенотипов в качестве обучающих данных, изучения порогового значения из обучающих данных, а затем классификации удерживаемого фенотипа путем вычисления его среднего сходства с каждым фенотипом обучающих данных в каждом классе или подмножество и классифицировать его как принадлежащее к любой категории, для которой среднее сходство с другими фенотипами в этой категории превышает изученный порог.Эффективность этой классификационной задачи с использованием каждой сети оценивалась с использованием метрики F 1 , где назначения функциональных категорий для каждого гена, о которых сообщали Ллойд и Мейнке (2012), считались правильными классификациями (таблица 3). Самые простые методы НЛП (набор слов и набор слов) превзошли Oellrich, Walls et al. (2015) вручную курировала сеть операторов EQ для этой задачи классификации во всех случаях, в то время как использование сгенерированных с помощью вычислений операторов EQ или встраивания документов, созданных с помощью Doc2Vec, только в некоторых случаях превосходило курируемую сеть операторов EQ.
Таблица 3 Оценка (F 1 баллов) для каждого метода, используемого для категоризации генов Arabidopsis по функциям.
В совокупности эти результаты показывают, что даже несмотря на то, что сети, созданные с помощью вычислений, значительно отличаются от сети, созданной вручную (рис. 3), они в целом одинаково хорошо или лучше справляются с задачами, связанными с организацией генов Arabidopsis в функциональные группы. . Затем мы исследовали, как эти сети сравниваются в задаче прогнозирования членства в биохимических путях для определенных генов как внутри одного вида, так и среди множества видов.
Вычислительные методы превосходят ручную обработку для восстановления генов, участвующих в биосинтезе антоцианов как внутри, так и между видами
Oellrich, Walls et al. (2015) проиллюстрировали полезность использования представлений фенотипов в выражениях EQ для предоставления семантической информации, необходимой для восстановления общего членства причинных генов в регуляторных и метаболических путях. В частности, они показали, что при запросе сети сходства фенотипов с шестью видами с геном c2 ( colorless2 ) кукурузы, который участвует в биосинтезе антоцианов, гены c1 , r1 и b1 ( colourless1 , red1 и booster1 ), которые также участвуют в биосинтезе антоцианов в кукурузе.Запрос в этом случае определяется как возвращение других генов в сети сходства, ранжированных с использованием максимального значения ребер, соединяющих фенотип, соответствующий гену запроса, и фенотип, соответствующий каждому другому гену в сети. В наборе данных 2747 генов, поэтому запрос с одним геном возвращает ранжированный список из 2746 генов. Этот результат был включен Oellrich, Walls et al. (2015) в качестве конкретного примера общей полезности сети фенотипического сходства для возврата других участников пути или регуляторной сети гена при запросе с одним геном.См. Рисунок 1 для общей иллюстрации этой концепции.
Чтобы оценить ту же полезность в сетях сходства фенотипов, которые мы создали с помощью вычислительных методов, и сравнить их полезность с полезностью сети от Oellrich, Walls et al. (2015), созданный с использованием составленных вручную утверждений EQ, мы сначала расширили набор генов пути антоцианов кукурузы, включив в него те, которые присутствуют в описании пути, приведенном Li et al. (2019) и перечислены в дополнительной таблице 1 этой публикации.Из этих генов 10 присутствуют в Oellrich, Walls et al. (2015) (таблица 4). Кроме того, мы аналогичным образом идентифицировали набор из генов Arabidopsis , которые, как известно, участвуют в биосинтезе антоцианов (перечислены в таблице 1 Appelhagen et al., 2014), которые присутствовали в Oellrich, Walls et al. (2015) набор данных. Это дало всего 16 генов Arabidopsis (таблица 5).
Таблица 4 Гены кукурузы, участвующие в биосинтезе антоцианов.
Таблица 5 Arabidopsis гена, участвующих в биосинтезе антоцианов.
Восстановление генов биосинтеза антоцианов
в пределах одного вида
Используя каждую сеть сходства фенотипов, каждый ген биосинтеза антоцианов одного вида итеративно использовался в качестве запроса к сети. Был определен ранг каждого другого гена в наборе генов биосинтеза антоцианов, соответствующих тому же виду, что и запрос. Мы сгруппировали ранги в интервалы шириной 10 для рангов меньше или равной 50 и объединили все ранги больше 50 в один интервал.Для каждой сети сходства фенотипов были рассчитаны среднее значение и стандартное отклонение количества генов биосинтеза антоцианов в каждой ячейке (рис. 5). Среднее количество генов, попавших в первую десятку по всем запросам, было больше для всех сетей, созданных с помощью вычислений, чем для сети, построенной из вручную подобранных операторов EQ, хотя разброс по запросам был высоким. В целом, вычислительные сети, построенные на основе предсказанных операторов EQ, лучше всего справились с этой задачей, тогда как сеть, построенная с использованием вручную подобранных EQ, показала худшие результаты.Сети, построенные с использованием числовых векторных представлений (набор слов, набор слов и Doc2Vec), были промежуточными по производительности как группа (рисунок 5).
Рис. 5 Ранжирование генов биосинтеза антоцианов кукурузы (A) или Arabidopsis (B) при запросе сетей сходства фенотипов, созданных с использованием генов одного и того же вида. Сети фенотипов организованы по методу, используемому для их создания ( столбца ), и по тому, были ли эти методы применены к описаниям фенотипа или фенотипа ( строки ).Значение ранга указывает диапазон ранжирования для каждого столбца на графиках (1–10, 11–20 и т. Д.), А величина ранга указывает среднее количество генов биосинтеза антоцианов, которые были ранжированы в заданном диапазоне по всем запросам. Планки ошибок указывают на одно стандартное отклонение ранговых величин в каждом диапазоне по всем запросам.
Восстановление генов биосинтеза антоциана между двумя видами
Чтобы определить, работают ли методы одинаково как внутри, так и между видами, мы повторили анализ, описанный в предыдущем разделе ( Восстановление генов биосинтеза антоциана в пределах одного вида ), но вместо количественной оценки ранги всех генов биосинтеза антоцианов от того же вида, что и запрашиваемый ген, мы количественно оценили ранги всех генов антоцианов, которые произошли от других видов.Другими словами, гена Arabidopsis, использовали для запроса генов кукурузы, а гены кукурузы использовали для запроса генов Arabidopsis . Как показано на рисунке 6, сеть сходства фенотипов, построенная из вручную подобранных утверждений EQ, не восстановила (предоставила ранги меньше или равные 50) ни один из генов биосинтеза антоцианов при запросе с генами других видов. Сети, созданные с использованием подходов набора слов и набора слов, или с помощью Doc2Vec, работали аналогично, восстанавливая в среднем менее одного гена биосинтеза антоцианов на запрос.Только сети, построенные на основе сгенерированных с помощью вычислений утверждений EQ, в среднем восстановили заметное количество генов биосинтеза антоцианов по запросам между видами (рис. 6).
Рис. 6 Ранжирование генов биосинтеза антоцианов кукурузы (A) или Arabidopsis (B) при запросе сетей сходства фенотипов, созданных с использованием генов других видов. Сети фенотипов организованы по методу, используемому для их создания ( столбца ), и по тому, были ли эти методы применены к описаниям фенотипа или фенотипа ( строки ).Значение ранга указывает диапазон ранжирования для каждого столбца на графиках (1–10, 11–20 и т. Д.), А величина ранга указывает среднее количество генов биосинтеза антоцианов, которые были ранжированы в заданном диапазоне по всем запросам. Планки ошибок указывают на одно стандартное отклонение ранговых величин в каждом диапазоне по всем запросам.
Обсуждение
Вычислительно созданные представления фенотипов полезны
Основная цель создания представлений фенотипов, которые легко вычислить (операторы EQ, векторные вложения и т. Д.)) заключается в построении сетей сходства, которые позволяют использовать один фенотип в качестве запроса для получения похожих фенотипов. Этот процесс служит средством обнаружения родства между фенотипами (потенциальными фенологами) внутри и между видами, генерируя, таким образом, гипотезы о лежащей в основе генетической родственности (обзор у Oellrich, Walls et al., 2015).
Было продемонстрировано, что вычислительные методы, обсуждаемые в этой работе, лишь частично воспроизводят сеть сходства фенотипов, построенную Oellrich, Walls et al.(2015) с использованием вручную подобранных операторов EQ ( Сравнение вычислительных сетей с вручную подобранной сетью ). Несмотря на ограниченное сходство между сетью, построенной из вручную отобранных аннотаций, и сетями, созданными с помощью вычислений, сети, созданные с помощью вычислений, работали так же или лучше, чем сеть, созданная вручную (на основе тщательно отобранных утверждений EQ) с точки зрения правильной организации фенотипов и их причинных причин. гены в функциональные категории на нескольких иерархических уровнях ( вычислительных методов превосходит ручное курирование для функциональной категоризации генов в Arabidopsis).Кроме того, каждая сеть, созданная с помощью вычислений, работала лучше, чем созданная вручную сеть для запросов к генам биосинтеза антоцианов кукурузы или Arabidopsis , чтобы вернуть другие гены биосинтеза антоцианов от того же вида ( Восстановление генов биосинтеза антоциана в пределах в пределах Species ), задача, первоначально использовавшаяся для демонстрации полезности сети фенотипического сходства, построенной Oellrich, Walls et al. (2015).
Более того, сети, построенные на основе вычислений сгенерированных операторов EQ, были полезны для повторного захвата генов биосинтеза антоцианов от видов, отличных от видов происхождения для запрашиваемой пары ген / фенотип.Ни одна из других сетей, включая сеть, построенную на основе тщательно подобранных операторов EQ, не предлагала эту утилиту для этой задачи ( Восстановление генов биосинтеза антоциана между двумя видами ). Этот конкретный результат указывает на то, что высокая точность построенных операторов EQ не требуется специально для таких задач, как поиск связанных генов у разных видов, потому что потенциально неточные (предсказанные вычислением) операторы EQ создали более успешную сеть для этой задачи. Воспроизведение этих анализов с описанием фенотипов в другой биологической области, например, у позвоночных, определило бы, будут ли эти результаты обобщаться на дополнительные группы видов и наборы данных.
Взятые вместе, эти результаты по этому конкретному набору данных описаний фенотипов предполагают, что, хотя утверждения EQ, сгенерированные путем ручного курирования, вероятно, являются наиболее точным и информативным вычислимым представлением данного фенотипа в конкретных случаях, другие представления, созданные полностью вычислительным способом без вмешательства человека. способны удовлетворить или превзойти производительность созданных вручную аннотаций для задач в масштабе набора данных, таких как сортировка фенотипов и генов по функциональным категориям, а также в случае конкретных задач, таких как запросы с определенными генами для восстановления других генов, участвующих в тот же путь.Следовательно, в случаях, когда объем данных велик, результаты считаются прогнозирующими, а ручное отслеживание нецелесообразно, использование автоматизированных методов аннотации для создания крупномасштабных сетей сходства фенотипов является стоящей целью и может предоставить биологически значимую информацию, которая может использоваться для генерации гипотез, включая предсказание нового гена-кандидата.
Полезны несколько подходов к представлению естественного языка
Аннотации операторов EQ, содержащие термины онтологии, обеспечивают взаимодействие с совместимыми аннотациями из различных источников данных.Они также являются удобочитаемым форматом аннотаций, что означает, что знающий человек может исправить некорректную аннотацию, выбрав более подходящий термин онтологии (процесс, который невозможен при использовании встраивания абстрактных векторов). Их единообразная структура также обеспечивает средства явного запроса фенотипов с участием биологического объекта, который похож на некоторую структуру или процесс (например, трихомы) или соответствует определенному качеству (например, увеличение физического размера). Аннотации, основанные на онтологиях, могут увеличить объем информации, связанной с фенотипом (путем вывода предковых терминов, которые специально не упоминаются в описании фенотипа), но не обязательно полностью отражают детали и семантику описания естественного языка.
По этой причине будущие представления фенотипов в реляционных базах данных с целью создания сетей сходства фенотипов в большом объеме фенотипов, описанных в литературе и в базах данных, вероятно, должны включать как основанные на онтологии аннотации, описывающие фенотипы, так и исходные описания на естественном языке. Хотя количество фенотипов в наборе данных, используемом здесь и описанном в Oellrich, Walls et al. (2015) относительно невелик, результаты этой работы указывают на полезность исходных текстовых представлений в качестве мощного средства вычисления сходства между фенотипами, особенно внутри одного вида.Вычислительно сгенерированные операторы EQ, которые в контексте этого исследования не часто соответствуют критериям полностью логически подобранных операторов EQ, оказались более полезными при любом другом подходе к восстановлению биологически связанных генов у разных видов.
Ансамблевые методы часто применяются в области машинного обучения, где для решения проблемы используются несколько методов, при этом модель более высокого уровня определяет, какой метод будет наиболее полезным при решении каждого нового экземпляра проблемы.Возможно, что такой подход может быть применен для измерения сходства между фенотипами для создания единой крупномасштабной сети, где значения сходства основаны на наилучшем возможном методе оценки текстовых представлений каждой пары конкретных фенотипов.
Дополнительные проблемы с представлением оператора EQ
Хотя термины онтологии и операторы EQ, составленные из терминов онтологии, представляют собой насыщенное информацией представление фенотипов и фенотипов, гибкость, в которой термины и утверждения могут представлять конкретный фенотип, может ограничивать возможность компьютерного распознавания биологическое сходство.Графические структуры самих онтологий, метрики, используемые для оценки семантического сходства, и неоднозначность, присущая как естественному языку, так и выражениям EQ-выражений фенотипов и фенотипов, потенциально могут способствовать возникновению этой проблемы.
В качестве одного из примеров в Oellrich, Walls et al. (2015), описание феномена «полная потеря цветков» было аннотировано оператором EQ, сущность которого — цветков , тогда как вычислительно идентифицированным объектом с использованием методов, описанных в этой работе, было цветков .В этом случае сходство Жаккара между этими двумя терминами онтологии составило 0,286, что для сравнения меньше, чем сходство Жаккара между формированием цветка и формированием листа в контексте графа онтологии. Этот выбранный пример иллюстрирует возможные расхождения между истинным биологическим сходством и семантическим сходством, измеренным с использованием показателей на основе графиков. Хотя каждая метрика семантического сходства вычисляет это значение по-разному, все те, которые используют иерархическую природу онтологии, ограничены структурой самого графа.
Различия в том, как люди и вычислительные методы интерпретируют то, как фенотип в целом следует концептуализировать, также могут создавать представления, скрывающие истинное сходство, измеряемое с помощью показателей на основе графиков. В другом примере от Oellrich, Walls et al. (2015), описание фена «тычинки, превращенные в пестики» было аннотировано двумя различными утверждениями EQ. Первый оператор EQ использует реляционное качество. имеет меньшее количество частей типа , чтобы указать на отсутствие тычинок в этом фенотипе, а второй использует реляционное качество. имеет дополнительные части типа , чтобы указать на присутствие пестиков в этом фенотипе.Такое представление фенотипа имеет логический смысл, но его нелегко создать с помощью вычислений, поскольку оно абстрактно описывает результат преобразования, который явно присутствует в описании естественного языка и отличается от представлений, сгенерированных с помощью вычислений, которые сосредоточены на явном содержании (т. Е. те, которые используют реляционное качество , преобразованы в ).
Наконец, в этом исследовании рассматривался набор данных, полностью состоящий из фенотипических описаний на английском языке, и возможность обобщения этих методов на другие языки не обсуждается.Определенно вероятно, что структурные различия между языками приведут к различиям в том, как работают определенные методы вычислений по сравнению с описаниями на этих языках, но такой анализ выходит за рамки данной работы.
Расширение этой работы на множество текстовых данных, доступных в базах данных и в литературе
Мы планируем применить методы семантической аннотации, вычисление семантического сходства на основе онтологий и вычисление семантического сходства на основе естественного языка к большому количеству текстовых данных доступны в существующих базах данных модельных организмов растений и в биологической литературе.Для последнего это потребует дополнительной задачи извлечения описаний фенотипов, а также генов, вызывающих эти фенотипы, в качестве отдельного этапа идентификации и обработки. Мы планируем использовать существующие работы в области распознавания именованных сущностей, специфичных для генов (Wei et al., 2015), и извлечения отношений, а также существующие методы извлечения информации, относящейся к фенотипам, например, разработанные с использованием векторных представлений фенотипа. описания (Xing et al., 2018) и представления описаний фенотипов в виде грамматического дерева (Collier et al., 2015). Поскольку размер применимого набора данных увеличивается с помощью этих средств, мы продолжим анализировать эффективность методов из областей машинного обучения и НЛП для построения биологически значимых сетей из этих фенотипических данных, включая дополнительные методы, которые не были включены в результаты. представлен здесь. Например, Sent2Vec (Pagliardini et al., 2018) — это еще один метод оценки сходства текста, который использует другой подход, чем Doc2Vec для встраивания текста в виде числовых векторов, и показал хорошие результаты при обучении на корпусах наук о жизни (Chen et al., 2018). Ожидается, что эти следующие шаги позволят исследователям начать вычисление описаний фенотипов напрямую и обеспечат многообещающее будущее для перспективных подходов к генетическим исследованиям, где фенотипы могут использоваться для предсказания новых генов-кандидатов так же легко, как поиск сходства последовательностей может использоваться для идентификации предполагаемых гомологов. из данных последовательности.
Заявление о доступности данных
Набор данных фенотипов и описаний фен, а также соответствующие подготовленные вручную утверждения EQ, используемые в этой работе, доступны в качестве дополнительных данных Oellrich, Walls et al.(2015). Иерархическая функциональная категоризация набора из генов Arabidopsis , использованная в этой работе, доступна в качестве дополнительных данных Lloyd and Meinke (2012). Код, использованный для получения результатов этой работы, доступен по адресу github.com/irbraun/phenologs. Файлы, необходимые для воспроизведения обсуждаемых результатов, наборы данных, используемые для создания рисунков, представленных в этой работе, и другие дополнительные файлы доступны по адресу doi.org/10.5281/zenodo.3255020. Это хранилище данных также включает версии ранее описанных наборов данных, доступных в качестве дополнительных данных Oellrich, Walls et al.(2015) и Lloyd and Meinke (2012), чтобы сделать это исследование воспроизводимым без каких-либо дополнительных внешних файлов.
Финансирование
В проведении этой работы авторов поддержали грант президентских междисциплинарных исследований Университета штата Айова, стипендиальная программа Института наук о растениях Университета штата Айова и стажировка NSF по прогнозированию феноменов растений (# DGE-1545453) .
Вклад авторов
IB и CL-D вместе внесли свой вклад в концепцию и дизайн исследования.IB систематизировал данные, провел анализ и написал рукопись. IB и CL-D внесли свой вклад в редактирование рукописи, а также прочитали и одобрили окончательную версию.
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта рукопись опубликована в виде препринта на doi.org/10.1101/689976. Мы благодарим Лизу Харпер, Sowmya Vajjala и Ramona Walls за полезные обсуждения и предложения.Мы признательны RCN по фенотипам онтологии NSF (# DBI-09) за создание основы для этой работы, объединив усилия биологов-биологов и компьютерных биологов для выработки общего словарного запаса, а также за их поддержку участникам пилотного проекта фенотипа растений, которые разработали методику Oellrich, Walls. и другие. (2015), на которые опирался наш анализ. Мы благодарим рецензентов за их ценные советы. На основании их предложений рукопись была значительно улучшена.
Сноски
- Что касается структуры предложения, сущность представляет подлежащее, а качество представляет предикат.Качества также в другом месте упоминаются как атрибуты, особенности или характеристики биологической структуры или процесса.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2019.01629/full#supplementary-material
Ссылки
Altenhoff, AM , Boeckmann, B., Capella-Gutierrez, S., Dalquen, DA, DeLuca, T., Forslund, K., et al. (2016). Стандартизированный бенчмаркинг в поисках ортологов. Нат. Методы 13, 425. doi: 10.1038 / nmeth4830
PubMed Реферат | CrossRef Полный текст | Google Scholar
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., Lipman, D. J. (1990). Базовый инструмент поиска локального выравнивания. J. Mol. Биол . 215, 403–410. DOI: 10.1016 / S0022-2836 (05) 80360-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Appelhagen, I., Thiedig, K., Nordholt, N., Schmidt, N., Huep, G., Sagasser, M., et al. (2014).Обновленная информация о прозрачных мутантах семенников из Arabidopsis thaliana : характеристика новых аллелей из изогенной коллекции. Планта 240 (5), 955–970. doi: 10.1007 / s00425-014-2088-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эшбернер М., Болл К. А., Блейк Дж. А., Ботстайн Д., Батлер Х., Черри Дж. М. и др. (2000). Генная онтология: инструмент для объединения биологии. Нат. Genet. 25 (1), 25–29. DOI: 10.1038 / 75556. Gen.arXiv: 10614036
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bada, M., Eckert, M., Evans, D., Garcia, K., Shipley, K., Sitnikov, D., et al. (2012). Аннотация концепции в корпусе КРАФТ. BMC Bioinf . 13, 161.doi: 10.1186 / 1471-2105-13-161
CrossRef Полный текст | Google Scholar
Браун, И., Балхофф, Дж. П., Берардини, Т. З., Купер, Л., Гкоутос, Г., Харпер, Л. и др. (2018). Вычислимые фенотипы позволяют проводить сравнительные и предсказательные феномены среди видов растений и во всех сферах жизни. Заявл. Семант. Technol. Биодайверы. Sci. 187–206.
Google Scholar
Чен, К., Ифань, П., Чжиюн, Л. (2018). BioSentVec: создание встраиваемых предложений для биомедицинских текстов. arXiv: 1810.09302 [cs.CL].
Google Scholar
Кольер, Н., Гроза, Т., Смедли, Д., Робинсон, П. Н., Оеллрих, А., Ребхольц-Шуманн, Д. (2015). PhenoMiner: от текста к базе данных фенотипов, связанных с заболеваниями OMIM. База данных 2015 (1), 1–12.doi: 10.1093 / database / bav104
CrossRef Полный текст | Google Scholar
Купер Л., Уоллс Р. Л., Эльзер Дж., Гандольфо М. А., Стивенсон Д. У., Смит Б. и др. (2013). Онтология растений как инструмент сравнительной анатомии растений и геномного анализа. Physiol растительных клеток. 54 (2), 1–23. DOI: 10.1093 / pcp / pcs163
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cui, H. (2012). CharaParser для детальной семантической аннотации морфологических описаний организмов. J. Am. Soc. Инф. Sci. Technol. 63 (4), 738–754. DOI: 10.1002 / asi. arXiv: 08031716
CrossRef Полный текст | Google Scholar
Cui, H., Dahdul, W., Dececchi, A. T., Ibrahim, N., Mabee, P., Balhoff, J. P., et al. (2015). CharaParser + EQ: оценка производительности без золотого стандарта ». Proc. Доц. Инф. Sci. Technol. 52 (1), 1–10. doi: 10.1002 / pra2.2015.145052010020
CrossRef Полный текст | Google Scholar
Dahdul, W., Manda, P., Cui, H., Balhoff, J. P., Dececchi, T. A., Ibrahim, N., et al. (2018). Аннотация фенотипов с использованием онтологий: золотой стандарт для обучения и оценки систем обработки естественного языка. База данных (Оксфорд). 2018, 1–17. doi: 10.1093 / database / bay110
CrossRef Полный текст | Google Scholar
Фальгрен, Н., Малия, А. Г., Иван, Б. (2015). Свет, камера, действие: высокопроизводительное фенотипирование растений 32 готово для крупного плана. Curr. Opin. Plant Biol. DOI: 10.1016 / j.pbi.2015.02.006
PubMed Реферат | CrossRef Полный текст | Google Scholar
Gehan, M.A., Fahlgren, N., Abbasi, A., Berry, J.C., Callen, S.T., Chavez, L., et al. (2017). PlantCV v2: программное обеспечение для анализа изображений для высокопроизводительного фенотипирования растений. ПирДж . 5, е4088. doi: 10.7717 / peerj.4088
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гкоутос, Г. В., Грин, Э. К. Дж., Мэллон, А.-м., Хэнкок, Дж. М., Дэвидсон, Д. (2005). Использование онтологий для описания фенотипов мышей. Genome Biol. 6 (1), R8. doi: 10.1186 / gb-2004-6-1-r8.R8
PubMed Реферат | CrossRef Полный текст | Google Scholar
Green, J. M., Appel, H., Rehrig, E. M., Harnsomburana, J., Chang, J.-F., Balint-Kurti, P., et al. (2012). PhenoPhyte: гибкий доступный метод количественной оценки 2D-фенотипов на основе изображений. Методы растений 8, 45. doi: 10.1186 / 1746-4811-8-45
PubMed Реферат | CrossRef Полный текст | Google Scholar
Hailu, N. D., Bada, M., Хадгу, А. Т., Хантер, Л. Э (2019). Распознавание биомедицинских концепций с использованием моделей глубоких нейронных последовательностей. bioRxiv . 1–21. doi: 10.1101 / 530337
CrossRef Полный текст | Google Scholar
Hastings, J., De Matos, P., Dekker, A., Ennis, M., Harsha, B., Kale, N., et al. (2013). Справочная база данных и онтология ChEBI для биологически релевантной химии: улучшения на 2013 г. . Nucleic Acids Res. 41 (D1), 456–463. doi: 10.1093 / nar / gks1146
CrossRef Полный текст | Google Scholar
Homma, N., Takei, Y., Tanaka, Y., Nakata, T., Terada, S., Kikkawa, M., et al. (2003). Белок суперсемейства кинезинов 2A (KIF2A) подавляет расширение коллатеральных ветвей. Ячейка 114, 229–239. DOI: 10.1016 / S0092-8674 (03) 00522-1
PubMed Реферат | CrossRef Полный текст | Google Scholar
Лау, Дж. Х., Болдуин, Т. (2016). Эмпирическая оценка doc2vec с практическим пониманием создания встраиваемых документов. arXiv: 1607.05368 [cs.CL].
Google Scholar
Le, Q.В., Миколов Т. (2014). Распределенные представления приговоров и документов. arXiv: 1405.4053 [cs.CL].
Google Scholar
Li, T., Zhang, W., Yang, H., Dong, Q., Ren, J., Fan, H., et al. (2019). Сравнительный анализ транскриптома выявляет по-разному экспрессируемые гены, связанные с тканеспецифическим накоплением антоцианов в околоплоднике и алейроновом слое кукурузы. Sci. Rep. 9, 1–12. doi: 10.1038 / s41598-018-37697-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ллойд, Дж., Мейнке, Д. (2012). Исчерпывающий набор данных генов с мутантным фенотипом потери функции в арабидопсисе . Plant Physiol. 158 (3), 1115–1129. DOI: 10.1104 / pp.111.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lu, L., Lee, Y.-R. Дж., Пан, Р., Малуф, Дж. Н., Лю, Б. (2004). Внутренний моторный кинезин связан с аппаратом Гольджи и играет роль в морфогенезе трихомов у arabidopsis . Мол. Биол. Ячейка 16, 811–823.doi: 10.1091 / mbc.e04-05-0400
PubMed Аннотация | CrossRef Полный текст | Google Scholar
МакГэри, К. Л., Парк, Т. Дж., Вудс, Дж. О., Ча, Х. Дж., Уоллингфорд, Дж. Б., Маркотт, Э. М. (2010). Систематическое открытие неочевидных моделей болезней человека через ортологичные фенотипы. Proc. Natl. Акад. Sci. 107 (14), 6544–6549. DOI: 10.1073 / pnas.00107. arXiv: 1408.1149
CrossRef Полный текст | Google Scholar
Миколов, Т., Чен, К., Коррадо, Г., Дин, Дж.(2013). Эффективное оценивание представлений слов в векторном пространстве. arXiv: 1301.3781 [cs.CL].
Google Scholar
Миллер, Н. Д., Хаас, Н. Дж., Ли, Дж., Кэпплер, С. М., де Леон, Н., Сполдинг, Э. П. (2017). Надежный и высокопроизводительный метод автоматического вычисления атрибутов початков кукурузы, початков и ядер кукурузы по изображениям. Плант Дж. 89, 169–178.doi: 10.1111 / tpj.13320
PubMed Реферат | CrossRef Полный текст | Google Scholar
Mungall, C.Дж., Гкоутос, Г. В., Смит, К. Л., Хендель, М. А., Льюис, С. Е., Эшбернер, М. (2010). Интеграция онтологий фенотипов для нескольких видов. Genome Biol. 11 (1), 1–16. doi: 10.1186 / gb-2010-11-1-r2
CrossRef Полный текст | Google Scholar
Musen, M.A., Noy, N.F., Shah, N.H., Whetzel, P.L., Chute, C.G., Story, M.A., et al. (2012). Национальный центр биомедицинской онтологии. J. Am. Med. Доцент информатики . doi: 10.1136 / amiajnl-2011-000523
CrossRef Полный текст | Google Scholar
Oellrich, A., Уоллс, Р. Л., Кэннон, Э. К., Кэннон, С. Б., Купер, Л., Гардинер, Дж. И др. (2015). Онтологический подход к сравнительной феномене растений. Растительные методы 11 (1), 1–15. doi: 10.1186 / s13007-015-0053-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пальярдини, М., Пракхар, Г., Мартин, Дж. (2018). Неконтролируемое обучение встраиванию предложений с использованием композиционных функций n-грамм, In: Proceedings of the North American Conference 2018 Chapter 77 of the Association for Computational Linguistics: Human Language Technologies Volume 1 (Long Papers) (Association for Computational Linguistics), 528– 540.doi: 10.18653 / v1 / n18-1049
CrossRef Полный текст | Google Scholar
Resnik, P. (1999). Семантическое сходство в таксономии: мера, основанная на информации, и ее применение к проблемам неоднозначности в естественном языке. J. Artif. Intell. Res. 11, 95–130.doi: 10.1613 / jair.514. arXiv: 1105.5444
CrossRef Полный текст | Google Scholar
Schla¨pfer, P., Zhang, P., Wang, C., Kim, T., Banf, M., Chae, L., et al. (2017). Полногеномное прогнозирование метаболических ферментов, путей и кластеров генов у растений. Plant Physiol. 173, 2041–2059. doi: 10.1104 / pp.16.01942.15
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарма М., Кортес-Крус М., Ахерн К. Р., МакМаллен М., Брутнелл Т. П., Чопра С. (2011). Идентификация продукта гена pr1 завершает путь биосинтеза антоцианов кукурузы. Генетика 188, 69–79. doi: 10.1534 / genetics.110.126136
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Slimani, T.(2013). Описание и оценка подходов к измерениям семантического сходства. Внутр. J. Comput. Заявление . 80, 25–33. doi: 10.5120 / 13897-1851
CrossRef Полный текст | Google Scholar
Тессен, А. Э., Хонг, К., Дмитрий, М. (2012). Применение обработки естественного языка в науке о биоразнообразии. Adv. Bioinf. 2012, 1–17. doi: 10.1155 / 2012/3
CrossRef Полный текст | Google Scholar
Цейтлин, Э., Митчелл, К., Леговски, Э., Корриган, Дж., Чаван, Г., Якобсон, Р. С. (2016). NOBLE — Гибкое распознавание концепций для крупномасштабной биомедицинской обработки естественного языка. BMC Bioinf. 17 (1), 32. doi: 10.1186 / s12859-015-0871-y
CrossRef Полный текст | Google Scholar
Wei, C.-H., Hung-Yu, K., Zhiyong, L. (2015). GNormPlus: интегративный подход к тегированию генов, семейств генов и белковых доменов. Biomed. Res. Int. 2015, 1–7. doi: 10.1155 / 2015/
0
CrossRef Полный текст | Google Scholar
Whetzel, P.Л., Ной, Н. Ф., Шах, Н. Х., Александер, П. Р., Ньюлас, К., Тудораш, Т. и др. (2011). BioPortal: расширенная функциональность через новые веб-службы от национального центра биомедицинской онтологии для доступа и использования онтологий в программных приложениях. Nucleic Acids Res . 39, W541 – W545. DOI: 10.1093 / nar / gkr469
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вудс, Дж. О., Сингх-Блом, У. М., Лоран, Дж. М., МакГэри, К. Л., Маркотт, Э. М. (2013).Прогнозирование ассоциаций ген-фенотип у людей, мышей и растений с использованием фенологов. BMC Bioinf. 14 (1), 203.
Google Scholar
Xing, W., Qi, J., Yuan, X., Li, L., Zhang, X., Fu, Y., et al. (2018). Конвейер извлечения ген-фенотипических отношений из биомедицинской литературы с использованием подхода репрезентативного обучения. Биоинформатика 34 (13), i386 – i394. doi: 10.1093 / биоинформатика / bty263
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Генотип-фенотипическая карта комплексных признаков дрожжей: основные параметры и роль естественного отбора | Молекулярная биология и эволюция
Абстрактные
Большинство фенотипических признаков контролируются многими генами, но общая картина генотип-фенотипической карты (GPM) отсутствует.Например, ни у одного вида мы не знаем в целом, сколько генов влияет на признак и насколько велики эти эффекты. Также неясно, в какой степени GPM формируются естественным отбором. Здесь мы решаем эти фундаментальные вопросы, используя обратные генетические данные 220 морфологических признаков у 4718 почкующихся штаммов дрожжей, у каждого из которых отсутствует несущественный ген. Мы показываем, что 1) доля генов, влияющих на признак, варьируется от <1% до> 30%, составляя в среднем 6%, 2) на большинство признаков влияет гораздо больше генов с малым эффектом, чем с генами с большим эффектом, и 3) среднее значение влияние всех несущественных генов на признак резко снижается по мере увеличения предполагаемой важности признака для приспособленности.Анализ 3116 признаков экспрессии дрожжевых генов в 754 штаммах с делецией генов показывает аналогичную картину. Эти находки иллюстрируют силу общегеномной обратной генетики в картировании генотип-фенотип, раскрывают огромный диапазон генетической сложности фенотипических признаков и предполагают, что GPM клеточных организмов сформирован естественным отбором на мутационную устойчивость.
Введение
Описание, понимание и использование взаимосвязи между генотипами и фенотипами или карты генотип-фенотип (GPM) являются основными целями генетики (Wagner and Zhang 2011).Поскольку большинство фенотипических признаков, включая те, которые имеют отношение к заболеваниям человека, контролируются множеством генов (Falconer and Mackay 1996) и поскольку большинство генов влияют на более чем один признак (Wang et al. 2010), GPM представляет собой плотную двудольную сеть генов и traits, где граница между геном и признаком указывает на то, что ген влияет на признак, а ширина грани представляет размер эффекта (Wang et al. 2010). Традиционно GPM конструируется с помощью прямой генетики, которая использует исследования сцепления или ассоциации для выявления генетических вариантов, лежащих в основе конкретных фенотипических вариаций среди особей одного и того же вида (Mackay et al.2009 г.). Из-за ограниченной мощности и эффективности таких анализов, GPM человека и большинства модельных организмов остаются весьма неполными и неинформативными (Mackay et al. 2009; Manolio et al. 2009). Например, недавний крупномасштабный анализ сцепления оценил количество генов, влияющих на каждый из 18 признаков дрожжей (Ehrenreich et al. 2010). Однако, поскольку в исследовании сравнивались только два штамма, были выявлены только те генетические варианты, которые вызывают фенотипические различия между этими двумя штаммами.Следовательно, неизвестно ни распределение количества генов, которые могут повлиять на признак, ни распределение размеров воздействия этих генов на признак. Оценка этих фундаментальных параметров GPM имеет жизненно важное значение, поскольку они влияют на то, насколько изменчивым является конкретный признак в популяции, определяют лучшую стратегию для выявления лежащих в основе генетических вариантов фенотипических вариаций и предсказывают, насколько устойчива и адаптируется популяция к окружающей среде. проблемы.
В отличие от прямой генетики, обратная генетика выявляет фенотипические различия между людьми с известными генетическими различиями.При усилении высокопроизводительного фенотипирования систематически генерируемых мутантов обратная генетика может быть эффективным подходом к GPM. Например, доля генов, которые влияют на каждый из 12 физиологических и поведенческих признаков у мыши Mus musculus , была оценена с использованием 250 линий нокаута генов (Flint and Mackay 2009) (рис. 1 a ). Аналогичные оценки были сделаны для восьми морфологических, физиологических и поведенческих признаков у плодовой мухи Drosophila melanogaster на основе мутагенеза вставки Р-элемента (Mackay 2010) (рис.1 b ). Эти и несколько других исследований (Winzeler et al. 1999; Ramani et al. 2012) показали, что доля генов, влияющих на признак, может достигать 10-40% всех генов в геноме. Но насколько общими являются эти результаты, неясно, потому что количество изученных признаков и видов невелико. Что касается распределения генных эффектов на признак, существуют две конкурирующие гипотезы. Модель бесконечно малых Мазера (Mather 1941; Mackay 2001) утверждает, что многочисленные локусы имеют малые и похожие эффекты, в то время как Робертсон (1967) утверждает, что распределение является приблизительно экспоненциальным, с несколькими локусами большого эффекта и множеством локусов малого эффекта.Распределение размера эффекта вставок Р-элемента на количество щетинок на брюшной и грудинно-плевральной щетинках у Drosophila подтверждает модель Робертсона (Lyman et al. 1996), но универсальность этого вывода неизвестна. Хотя передовые генетические исследования нескольких видов также подтверждают модель Робертсона, окончательные выводы затрудняются из-за присущих этому методу предубеждений и ограничений (Mackay 2001).
Рис. 1.
Доля генов, влияющих на признак, с указанием среднего и медианного значений.( a ) Паттерны, возникающие из 12 признаков, исследованных на 250 линиях мышей с нокаутом. ( b ) Паттерны, возникающие из восьми признаков, исследованных в различных линиях вставки элементов P у плодовых мушек. В ( a ) и ( b ) каждая стрелка представляет одну черту. ( c ) Частотное распределение доли генов, влияющих на признак, полученное из 220 морфологических признаков, исследованных в 4718 линиях с делецией несущественных генов дрожжей.
Рис.1.
Доля генов, влияющих на признак, с указанием среднего и медианного значений. ( a ) Паттерны, возникающие из 12 признаков, исследованных на 250 линиях мышей с нокаутом. ( b ) Паттерны, возникающие из восьми признаков, исследованных в различных линиях вставки элементов P у плодовых мушек. В ( a ) и ( b ) каждая стрелка представляет одну черту. ( c ) Частотное распределение доли генов, влияющих на признак, полученное из 220 морфологических признаков, исследованных в 4718 линиях с делецией несущественных генов дрожжей.
Более глубокий вопрос о GPM — почему он выглядит именно так. В принципе, GPM может развиваться при мутации, дрейфе и отборе, но относительный вклад этих сил неуловим. Waddington и другие предположили, что GPM был сформирован естественным отбором на мутационную устойчивость, что привело к генетической канализации (Waddington 1942; de Visser et al. 2003). Точно так же естественный отбор мог привести к устойчивости организма к возмущениям окружающей среды или к ее канализации.Эти две формы канализации, если они верны, объяснили бы удивительную устойчивость живых организмов к генетическим и экологическим нарушениям, которые довольно распространены в природе (Scharloo 1991; Flatt 2005; Wagner 2005b; Alon 2007). Они также влияют на то, насколько популяция адаптируется и эволюционирует перед лицом мутаций и изменений окружающей среды (Гибсон и Вагнер, 2000; де Виссер и др., 2003; Вагнер, 2005b; Драги и др., 2010). Хотя отбор на устойчивость к окружающей среде общепризнан (Гибсон и Вагнер 2000), прямой отбор на генетическую устойчивость вызывает споры (Гибсон и Вагнер 2000; де Виссер и др.2003), за исключением случаев, когда частота вредных мутаций чрезвычайно высока и / или размер популяции огромен (например, у вирусов) (Wilke et al. 2001; Ciliberti et al. 2007; Sanjuan et al. 2007), поскольку для клеточных организмов такие ожидается, что отбор будет слабым (Wagner et al. 1997; Gibson and Wagner 2000), а предыдущие тесты с относительно небольшими данными дали неоднозначные результаты (Stearns and Kawecki 1994; Stearns et al. 1995; Houle 1998; Gibson and Wagner 2000; de Visser et al.2003; Proulx et al.2007). По-видимому, для его критической оценки необходимы более крупные и точные данные.
Чтобы ответить на эти фундаментальные вопросы об основных параметрах GPM и роли естественного отбора в формировании GPM, мы используем почкующиеся дрожжи Saccharomyces cerevisiae , у которых 220 морфологических признаков были количественно измерены путем анализа флуоресцентных микроскопических изображений трехкратно окрашенные клетки штамма дикого типа и 4718 мутантных штаммов, каждый из которых лишен несущественного гена (Ohya et al. 2005). Общность результатов морфологических признаков затем подтверждается путем анализа 3116 признаков экспрессии генов в штаммах дикого типа и 754 генных штаммах S.cerevisiae .
Результаты
Фракция генов, влияющих на морфологический признак
В эксперименте по фенотипированию дрожжей 220 морфологических признаков были измерены в нескольких клетках дикого типа из каждой из 123 репликативных популяций (Ohya et al. 2005). Кроме того, для каждого из 4718 мутантных штаммов было измерено несколько изогенных клеток из одной популяции по 220 признакам (Ohya et al. 2005). Чтобы определить, влияет ли удаление гена на признак, мы использовали тест Манна-Уитни U для сравнения значений признаков для нескольких клеток штамма с делецией гена и клеток дикого типа из произвольной реплицируемой популяции.Предварительно считается, что делеция гена влияет на признак, если значение P ниже 0,05. Распределение доли генов, влияющих на признак ( f mt ), показано на дополнительном рисунке S1a, Дополнительный материал онлайн. Среднее и медианное значение f mt составляют 0,37 и 0,38 соответственно. Чтобы исключить смешивающий фактор потенциальных различий в окружающей среде между мутантными штаммами и штаммами дикого типа в эксперименте и контролировать многократное тестирование, для каждого признака мы оценили фракцию ( f wt ) других 122 штаммов дикого типа. популяции, в которых значение признака значительно отличается от значения произвольной использованной популяции дикого типа.Мы обнаружили, что f wt является существенным (дополнительный рис. S1b, дополнительные материалы онлайн). Вычитая f wt из f mt , мы получили f генов , истинную долю генов, которые при удалении существенно влияют на признак. Мы обнаружили, что f гены сильно различаются по признакам (рис.1 c ) и что эта вариация значительно превышает случайное ожидание при гомогенных f генах ( P <0.01; перестановочный тест). В частности, на 37,7%, 39,6%, 20,0% и 2,7% признаков влияют <1%, 1–10%, 10–30% и> 30% всех несущественных генов в геноме дрожжей, соответственно. Среднее значение и медиана для генов f составляют 0,06 и 0,04 соответственно (рис. 1 c ). Эти результаты остаются аналогичными независимо от используемого порогового значения P- (дополнительный рис. S1c – d, дополнительный материал онлайн). Использование другой произвольной репликационной популяции штамма дикого типа дало аналогичные результаты.Для каждого признака мы также оценили f генов , исследуя, будет ли среднее значение признака мутанта выбросом в распределении средних значений 123 репликативных популяций дикого типа, но результаты были аналогичными (дополнительный рис. .S1e, Дополнительные материалы на сайте). Поскольку некоторые из 220 признаков сильно коррелированы, мы удалили те признаки, коэффициенты генетической корреляции которых превышают 0,7, в результате чего был получен набор данных с 54 признаками. Но средние и медианные значения f генов практически не изменились (дополнительный рис.S2, Дополнительные материалы онлайн).
Возможность обнаружения генного эффекта зависит от статистической мощности эксперимента, которая определяется точностью фенотипического измерения, размером выборки и постоянством среды, в которой фенотипируются различные штаммы. В данном случае количество измеренных клеток варьировалось в зависимости от признаков и штаммов. В среднем 91 и 95 клеток были измерены в штаммах дикого типа (на реплицитную популяцию) и делеционных штаммах, соответственно.Эффект изменчивости окружающей среды четко прослеживается в 123 популяциях дикого типа, поскольку стандартное отклонение среднего фенотипического значения среди 123 популяций в среднем в 2,48 раза больше средней ошибки цепочки, рассчитанной для отдельных популяций (дополнительный рисунок S3, дополнительный материал онлайн). Это наблюдение предполагает, что колебания окружающей среды, а не размер выборки или ошибка измерения являются доминирующим фактором, ограничивающим обнаружение генных эффектов в настоящем исследовании.
Средний размер эффекта делеции гена на морфологический признак
Мы определяем размер необработанного эффекта (ES) удаления гена на признаке как разницу между средним значением признака делеционного штамма и средним значением средних значений признака 123 реплицируемых популяций штамма дикого типа, разделенных по среднему значению средних значений признаков для 123 популяций дикого типа. Кумулятивное распределение вероятностей | ES |, или абсолютное значение ES, для 4718 генов по каждому из 220 признаков изображено кривой на рисунке 2 a .Это распределение показывает, что в большинстве случаев на признак влияет больше генов с малым воздействием, чем с генами с большим воздействием, как было предложено Робертсоном (1967). Рассмотрение только статистически значимых генных эффектов не меняет этого вывода. Среднее | ES | всех несущественных генов признака существенно различается по признаку со средним значением 0,098 (рис. 2 b ).
Рис. 2.
Распределение абсолютных величин чистых и чистых эффектов (| ES |) 4718 делеций несущественных генов по 220 морфологическим признакам у дрожжей.( a ) Кумулятивные распределения вероятностей исходных | ES | 4718 делеций генов по 220 признакам. Каждая кривая представляет признак и окрашена в соответствии с важностью признака (TI). Распределения показаны только в диапазоне 0 <| ES | <1, чтобы лучше различать разные кривые. ( b ) Распределение среднего необработанного | ES | среди 220 черт. ( c ) Кумулятивные распределения вероятностей net | ES | 4718 делеций генов по 220 признакам. ( d ) Распределение среднего нетто | ES | среди 220 черт.( e ) Распределение фенотипической изменчивости (CV) дикого типа среди 220 признаков.
Рис. 2.
Распределение абсолютных величин чистых и чистых эффектов (| ES |) 4718 делеций несущественных генов по 220 морфологическим признакам у дрожжей. ( a ) Кумулятивные распределения вероятностей исходных | ES | 4718 делеций генов по 220 признакам. Каждая кривая представляет признак и окрашена в соответствии с важностью признака (TI). Распределения показаны только в диапазоне 0 <| ES | <1, чтобы лучше различать разные кривые.( b ) Распределение среднего необработанного | ES | среди 220 черт. ( c ) Кумулятивные распределения вероятностей net | ES | 4718 делеций генов по 220 признакам. ( d ) Распределение среднего нетто | ES | среди 220 черт. ( e ) Распределение фенотипической изменчивости (CV) дикого типа среди 220 признаков.
Из-за неизбежных колебаний окружающей среды среди популяций клеток, которые были фенотипированы, мы вычислили net | ES | путем вычитания из сырых | ES | термин, называемый псевдо | ES |, который представляет собой абсолютную ES, ожидаемую от изменений окружающей среды и ошибки выборки, возникающей из-за ограниченного размера выборки (см. Материалы и методы).Кумулятивное распределение вероятностей сети | ES | 4718 генов по каждому из 220 признаков изображено кривой на рисунке 2 c . Опять же, это распределение поддерживает модель Робертсона (1967). Средняя чистая | ES | всех несущественных генов признака также существенно различается по признаку, со средним значением 0,035 (рис. 2 d ).
Для каждого признака фенотипические вариации среди изогенных клеток включают вариации, возникающие из-за стохастического шума признака, случайной ошибки измерения и изменчивости окружающей среды.Мы количественно оценили фенотипические вариации среди изогенных клеток дикого типа с помощью коэффициента вариации (CV = стандартное отклонение / среднее значение), включая как вариации среди клеток в популяции, так и вариации среди реплицированных популяций (см. Материалы и методы). Среднее значение CV 220 исследованных признаков составляет 0,41 (рис. 2 и ).
Более важные морфологические признаки более устойчивы к различным возмущениям
После описания основных параметров GPM для морфологии дрожжей мы исследуем потенциальную роль естественного отбора в формировании GPM.Гипотеза естественного отбора на устойчивость к окружающей среде предсказывает, что признаки, которые более важны для выживания и воспроизводства организма, имеют меньшие CV , , потому что естественный отбор на экологическую устойчивость признака усиливается с увеличением важности признака. Точно так же гипотеза естественного отбора в отношении генетической устойчивости предсказывает, что при определенных условиях признаки, которые более важны для выживания и воспроизводства организма, имеют меньшую чистую | ES |, поскольку такой GPM сводит к минимуму вредные эффекты случайных мутаций (Wagner et al.1997) (см. Материалы и методы). Чтобы проверить эти гипотезы, мы определяем важность признака (TI) в 100 раз меньшую приспособленность, вызванную 1% -ным изменением фенотипической ценности рассматриваемого признака, и оценили его с помощью нетто | ES | оценки и значения приспособленности штаммов с делецией гена в среде, где были собраны морфологические данные (Qian et al. 2012) (см. Материалы и методы). При оценке TI мы использовали 2779 штаммов с делецией генов, значения приспособленности которых относительно дикого типа меньше 1 (см. Материалы и методы).В результате 210 признаков (из 220) имеют TI> 0, 197 из которых значительно превышают 0 (номинальное значение P <0,05). Эти 210 признаков подлежали дальнейшему анализу.
Мы обнаружили, что CV признака уменьшается с ростом TI ( ρ = -0,692, P <10 −300 ; рис. 3 a ). Поскольку для оценки TI использовались фенотипические измерения дикого типа и мутантов, корреляция между CV и TI могла быть артефактической. Чтобы исключить эту возможность, мы оценили 1000 наборов значений псевдо-TI путем случайного перемешивания значений пригодности среди штаммов с делецией гена.В каждом наборе отрицательные значения TI игнорируются, потому что они не имеют биологического смысла. Мы рассчитали 1000 ранговых корреляций между CV и 1000 наборами положительного псевдо-TI. Поскольку эти ранговые корреляции нельзя напрямую сопоставить из-за разных размеров выборки, мы преобразовали корреляции () в оценки Фишера z на . Мы обнаружили, что истинный балл z (преобразованный из ρ = -0,692) был более отрицательным, чем все 1000 псевдо z баллов ( P <0.001; инжир. 3 b ), предполагая, что отрицательная корреляция между CV и TI является подлинной. Поскольку при расчете истинных баллов z и псевдо z использовались одни и те же фенотипические данные, их несоответствие не может быть вызвано ошибками измерения при фенотипировании. Скорее, он выявляет меньший стохастический шум и изменение окружающей среды для более важных признаков, что согласуется с гипотезой о том, что естественный отбор повысил фенотипическую устойчивость организмов к стохастическому шуму (Batada and Hurst 2007; Lehner 2008; Wang and Zhang 2011) и возмущениям окружающей среды ( Гибсон и Вагнер 2000).
Рис. 3.
Экологическая / стохастическая устойчивость и генетическая устойчивость морфологических признаков дрожжей. ( a ) CV признака среди изогенных клеток дикого типа снижается с повышением TI. Каждая точка — это особенность. ( b ) Распределение значений z Фишера, полученное на основе ранговой корреляции между CV и псевдо TI. ( c ) Средняя чистая | ES | Количество делеций гена на признаке уменьшается с увеличением TI, демонстрируя генетическую устойчивость.Каждая точка — это особенность. ( d ) Распределение Fisher z , полученное на основе ранговой корреляции между средними чистыми | ES | и псевдо-TI. ( e ) Распределение Fisher z , полученное из частичной ранговой корреляции между CV и псевдо TI, после контроля среднего чистого | ES |. ( f ) Распределение Fisher z , полученное из частичной ранговой корреляции между средними чистыми | ES | и псевдо-TI после контроля CV. В ( b ), ( d ), ( e ) и ( f ) реальный z , наблюдаемый по фактическим данным, обозначен стрелкой, а значение P — это вероятность того, что случайно выбранный псевдо z более отрицательный, чем реальный z .
Рис. 3.
Экологическая / стохастическая устойчивость и генетическая устойчивость морфологических признаков дрожжей. ( a ) CV признака среди изогенных клеток дикого типа снижается с повышением TI. Каждая точка — это особенность. ( b ) Распределение значений z Фишера, полученное на основе ранговой корреляции между CV и псевдо TI. ( c ) Средняя чистая | ES | Количество делеций гена на признаке уменьшается с увеличением TI, демонстрируя генетическую устойчивость. Каждая точка — это особенность.( d ) Распределение Fisher z , полученное на основе ранговой корреляции между средними чистыми | ES | и псевдо-TI. ( e ) Распределение Fisher z , полученное из частичной ранговой корреляции между CV и псевдо TI, после контроля среднего чистого | ES |. ( f ) Распределение Fisher z , полученное из частичной ранговой корреляции между средними чистыми | ES | и псевдо-TI после контроля CV. В ( b ), ( d ), ( e ) и ( f ) реальный z , наблюдаемый по фактическим данным, обозначен стрелкой, а значение P — это вероятность того, что случайно выбранный псевдо z более отрицательный, чем реальный z .
Мы также наблюдали отрицательную корреляцию между средним чистым | ES | по всем линиям делеций и TI (? = -0,793, P <10 -300 ; рис.3 c ), что указывает на то, что средний размер эффекта всех несущественных генов на признак уменьшается по мере того, как признак становится что еще более важно, поддерживая гипотезу естественного отбора на мутационную устойчивость. Этот результат был подтвержден путем сравнения наблюдаемого z (преобразованного из ρ = -0.793) с 1000 псевдо z баллов, преобразованных из корреляций между средним чистым | ES | и 1000 наборов положительного псевдо-TI ( P <0,001; рис. 3 d ).
Интересно, что мы не обнаружили значимой корреляции между TI признака и количеством генов, влияющих на признак ( f генов ) (смоделировано P = 0,07; дополнительный рисунок S4, дополнительные материалы онлайн). Следовательно, нижнее среднее чистое | ES | важных черт не потому, что меньше генов, влияющих на важные черты, а потому, что индивидуальные воздействия меньше.
Поскольку генетическая устойчивость может быть побочным продуктом естественного отбора для экологической / стохастической устойчивости или наоборот (гипотеза конгруэнтности) (Гибсон и Вагнер, 2000; де Виссер и др., 2003), важно проверить, соответствуют ли эти два типа устойчивости имеют независимое происхождение. Мы обнаружили, что частичная корреляция между CV и TI после контроля среднего чистого | ES | составляет ρ = −0,532 ( P = 1,1 × 10 −16 ), а частичная корреляция между средним чистым значением | ES | а TI после контроля CV составляет ? = -0.700 ( P = 4,7 × 10 −32 ). Следовательно, экологическая / стохастическая устойчивость и генетическая устойчивость не полностью связаны друг с другом и должны иметь свое собственное происхождение. Эти результаты были дополнительно подтверждены сравнением со случайными ожиданиями от 1000 наборов псевдо-TI (рис. 3 e и f ). Из-за потенциальной разницы в эффектах приспособленности положительных и отрицательных генных эффектов на признак, мы также повторно проанализировали данные, используя только положительные (или отрицательные) эффекты, но обнаружили, что результаты не изменились качественно (дополнительная таблица S1, дополнительные материалы онлайн; см. Материалы и методы).
Чтобы подтвердить, что значимые корреляции между CV, средним чистым | ES | и TI не связаны с высокими генетическими корреляциями между некоторыми признаками, мы использовали два подхода для получения менее коррелированных признаков. Во-первых, мы удалили сильно коррелированные признаки, как это было сделано для дополнительного рисунка S2, но отрицательную корреляцию между CV и TI и между средним чистым | ES | и TI все еще существуют, так же как и частичные корреляции (дополнительный рис. S5, дополнительные материалы онлайн). Во-вторых, мы выполнили анализ главных компонент с использованием сети | ES | матрица (см. Материалы и методы).Используя признаки главных компонентов, мы подтвердили отрицательную корреляцию между средним чистым | ES | и TI (дополнительный рис. S6, дополнительные материалы в Интернете).
Отрицательная корреляция между средним чистым | ES | и TI могут означать снижение чистой | ES | на важные черты или увеличение чистой | ES | за неважные черты характера; только первые поддерживают естественный отбор на генетическую устойчивость. Чтобы различить эти две возможности, мы проанализировали 4718 генов отдельно. Для каждого гена мы оценили ранговую корреляцию ( ρ TI- | ES | ) между важностью признака и чистой | ES | гена по признаку среди 210 признаков с предполагаемым TI.Найдено ρ TI- | ES | сильно различаются среди генов, хотя большинство (64,8%) генов имеют отрицательные значения ρ TI- | ES | значений (рис.4 а ). Мы разделили 210 черт пополам на группу менее важных и группу более важных черт. Затем мы, соответственно, вычислили среднее чистое | ES | 20% наиболее устойчивых генов (т.е. с наиболее отрицательными значениями ρ TI- | ES | ) и 20% наименее устойчивых генов (т.е. с наименьшими | ρ TI- | ES | | ценностей) по каждой группе признаков.Естественный отбор на мутационную устойчивость должен усиливаться по более важным признакам. Таким образом, для группы менее важных признаков мы ожидаем net | ES | быть похожими между наименее надежными и наиболее устойчивыми генами; но для группы более важных признаков net | ES | должен быть меньше для наиболее устойчивых генов, чем для наименее устойчивых генов. Противоположный образец несовместим с отбором на мутационную устойчивость. Мы обнаружили, что для менее важных признаков средняя чистая | ES | сходно между наиболее устойчивыми и наименее устойчивыми генами ( P = 0.21 — знаковый ранговый тест Уилкоксона; инжир. 4 б ). Но для более важных признаков средняя чистая | ES | из наиболее устойчивых генов становится значительно меньше, чем у наименее устойчивых генов ( P = 3,8 × 10 −22 , критерий знакового ранга Вилкоксона; рис. 4 b ). Эти результаты показывают, что отрицательная корреляция между TI и средним чистым | ES | вызвано уменьшением средней чистой | ES | значительной части генов по важным признакам, поддерживающим естественный отбор на генетическую устойчивость.
Рис. 4.
Вариация между генами по вкладу в генетическую устойчивость. ( a ) Частотное распределение ранговой корреляции гена ( ρ ) между его абсолютным чистым влиянием (| ES |) на признак и TI. Большинство генов показывают отрицательную корреляцию. ( b ) Влияние различий в величине между 20% наиболее устойчивыми (имеющими самые отрицательные значения ρ в a ) и 20% наименее устойчивыми (имеющими наименьшие | ρ | значения) генами на признаки разной важности. .Признаки делятся на две ячейки равного размера на основе TI: менее важные признаки и более важные признаки. На прямоугольной диаграмме (см. Шкалу на оси y ) нижний и верхний край прямоугольника представляют 25% квартиль ( q 1 ) и 75% квартиль ( q 3). ) соответственно. Горизонтальная линия внутри рамки указывает медианное значение (md). Усы простираются до самых крайних значений внутри внутренних ограждений, md ± 1,5 ( q 3 — q 1 ).Значения за пределами внутренних ограждений (выбросы) отмечены знаком плюса.
Рис. 4.
Вариация между генами влияет на генетическую устойчивость. ( a ) Частотное распределение ранговой корреляции гена ( ρ ) между его абсолютным чистым влиянием (| ES |) на признак и TI. Большинство генов показывают отрицательную корреляцию. ( b ) Влияние различий в величине между 20% наиболее устойчивыми (имеющими самые отрицательные значения ρ в a ) и 20% наименее устойчивыми (имеющими наименьшие | ρ | значения) генами на признаки разной важности. .Признаки делятся на две ячейки равного размера на основе TI: менее важные признаки и более важные признаки. На прямоугольной диаграмме (см. Шкалу на оси y ) нижний и верхний край прямоугольника представляют 25% квартиль ( q 1 ) и 75% квартиль ( q 3). ) соответственно. Горизонтальная линия внутри рамки указывает медианное значение (md). Усы простираются до самых крайних значений внутри внутренних ограждений, md ± 1,5 ( q 3 — q 1 ).Значения за пределами внутренних ограждений (выбросы) отмечены знаком плюса.
Фитнес-преимущество генетической устойчивости
Одна из основных причин, почему естественный отбор на предмет генетической устойчивости вызывает споры для клеточных организмов, заключается в том, что его коэффициент отбора, как ожидается, будет небольшим (Гибсон и Вагнер, 2000). Ниже мы покажем, что коэффициент отбора в данном случае достаточно велик, чтобы эффект естественного отбора превзошел эффект генетического дрейфа.
Если рассматривать только нулевые мутации, но не другие вредные мутации, преимущество пригодности ( g ) модификатора устойчивости будет равно ∑ ( μ i Δ s i / s i ) , где μ i — частота нулевых мутаций в гене i и составляет в среднем 2,15 × 10 −6 на ген на поколение у дрожжей (см. Материалы и методы), s i и s i — Δ s i — коэффициенты отбора против нулевой мутации гена i в отсутствие и в присутствии модификатора, соответственно, а ∑ указывает на суммирование по всем рассматриваемым генам (см. Материалы и методы. ).Модификатор строго выбран, когда S = 2 N e g = 2 × 10 7 × 2,15 × 10 −6 × ∑ (Δ s i / s i ) = 43∑ (Δ s i / s i ) значительно превышает 1, где N e — эффективный размер популяции и составляет ∼10 7 дюймов. дрожжи (Wagner 2005a). Например, g = 2,15 × 10 −5 и S = 430, если модификатор буферизует нулевые мутации 20 генов со средним значением Δ s i / s i = 0.5.
С учетом вышеизложенного, давайте оценим преимущество приспособляемости дрожжей, обусловленное наблюдаемой генетической устойчивостью. Это преимущество можно разделить на две части: 1) уменьшение средней величины эффекта всех генов по всем признакам и 2) большее уменьшение средней величины эффекта по более важным признакам. В идеале мы должны сравнивать приспособленность организма, полученную в результате реального GPM при наличии мутации, и приспособленность, полученную в результате изначального GPM, в котором величина эффекта не была уменьшена путем отбора.Это сравнение, однако, невозможно из-за трудности определения размеров наследственного эффекта. Вместо этого мы оценили приспособленность, полученную в результате гипотетического GPM, в котором размеры эффекта гена на различные признаки выбираются случайным образом (без замены) из наблюдаемых чистых размеров воздействия гена на эти признаки. Эта процедура дает консервативную оценку преимущества генетической устойчивости в пригодности, поскольку оценивается только часть 2). Используя этот подход, мы создали 1000 гипотетических GPM.
Мы построили многомерную линейную модель, в которой значения пригодности 2779 штаммов с делецией генов, которые менее подходят, чем дикий тип, объясняются фенотипами 220 признаков (см. Материалы и методы). Эта модель объясняет 45% различий в пригодности для используемых делеционных штаммов. Используя эту модель и GPM, мы можем предсказать пригодность при делеции гена. Например, прогнозируемая ожидаемая пригодность после делеции одного из 2779 несущественных генов из реального GPM равна 0.9443, что по существу идентично экспериментально определенной средней приспособленности (отличается на 4 × 10 −16 ) 2779 штаммов с делецией несущественных генов. Для 1000 гипотетических GPM прогнозируется, что приспособленность упадет до 0,8956 со стандартным отклонением 0,0091, когда случайно выбранный несущественный ген будет удален. Таким образом, пагубный эффект удаления среднего несущественного гена из реального GPM был уменьшен на впечатляющую долю Δ s / s = (0.9443 — 0,8956) / (1 — 0,8956) = 46,6% по сравнению с GPM с рандомизированной величиной эффекта. Следовательно, приспособленность дрожжей после делеции гена увеличилась на (0,9443 — 0,8956) / 0,8956 = 5,4%. Общее улучшение приспособленности, обусловленное устойчивостью к нулевым мутациям, составляет G = ∑ ( μ i Δ s i / s i ) = 2779 × 2,15 × 10 −6 × 46,6% = 2,8 × 10 −3 . Здесь мы используем G вместо g для обозначения комбинированного эффекта нескольких модификаторов.Поскольку вредные мутации, которые не полностью отменяют функцию гена, не учитывались в приведенном выше расчете, общий выигрыш в приспособляемости от мутационной устойчивости должен быть больше 2,8 × 10 -3 . Конечно, по мере того, как вредные мутации становятся менее серьезными после канализации, их равновесная частота в популяции будет увеличиваться. Следовательно, средняя приспособленность популяции вернется к прежнему значению (Wagner et al. 1997).
Генетическая устойчивость признаков экспрессии генов
Чтобы проверить, можно ли распространить генетическую устойчивость, наблюдаемую по морфологическим признакам, на другие признаки, мы обратимся к другому большому набору признаков, в котором уровень экспрессии каждого из 3116 генов дрожжей считается признаком.Генетическая устойчивость признаков экспрессии дрожжей ранее оценивалась путем расчета того, насколько экспрессия каждого гена варьируется между линиями с делециями гена, и тестирования, коррелирует ли степень вариации с важностью гена, но результаты были неоднозначными (Proulx et al. 2007). Мы расширили анализ с рассмотрения эффектов 276 делеций генов (Proulx et al. 2007) до 754, объединив несколько экспериментов с микрочипами, выполненных в богатой среде (см. Материалы и методы). Мы определяем необработанный ES удаляющего гена 1 на уровне экспрессии гена 2 по разнице в уровне экспрессии гена 2 между делеционным штаммом и штаммом дикого типа, деленной на уровень экспрессии гена 2 в диком типе.Поскольку уровни экспрессии были измерены в нескольких различных экспериментах с микрочипами, трудно оценить, является ли эффект статистически значимым. Но если использовать raw | ES | 0,2, 0,3, 0,4 и 0,5 в качестве потенциальных отсечений, средняя доля делеций гена, которые влияют на уровень экспрессии гена, составляет 20%, 10%, 5,5% и 3,3% соответственно. Таким образом, экспрессионные признаки сопоставимы по сложности с 220 морфологическими признаками.
В принципе, мы должны оценивать важность признаков экспрессии генов с помощью подхода, используемого для оценки TI морфологических признаков.Однако для этого потребуется прямое сравнение уровней экспрессии генов по разным данным микрочипа, что вряд ли будет надежным. Вместо этого мы последовали предыдущему исследованию (Proulx et al. 2007), чтобы использовать эффект приспособленности удаления гена в качестве заместителя для TI признака экспрессии гена. То есть экспрессия гена более важна, если эффект приспособленности от удаления гена больше. Этот прокси для TI является разумным, потому что эффект приспособленности, вызванный небольшим изменением экспрессии гена, сильно коррелирует с эффектом, вызванным удалением гена (Wang and Zhang 2011).Мы обнаружили, что независимо от того, измеряется ли TI по важности гена (т. Е. Категориальной) или по эффекту приспособленности от делеции гена (т. Е. Непрерывно), существует значимая отрицательная корреляция между важностью признака (TI) и средним абсолютным значением. ES делеции гена по признаку (| ES G |, индекс G указывает на генетическое нарушение) (таблица 1). Здесь ES G определяется так же, как ES для морфологических признаков. Мы также обнаружили отрицательную корреляцию между TI и средней абсолютной ES изменений окружающей среды (| ES E |, индекс E указывает на нарушение окружающей среды; см. Материалы и методы).Подобно результатам для морфологических признаков, корреляция между TI и | ES G | остается значимым после контроля | ES E | и корреляции между TI и | ES E | остается значительным после контроля | ES G | (таблица 1), предполагая, что ни генетическая, ни экологическая устойчивость экспрессии гена полностью не вызваны другим. Аналогичные результаты были получены при контроле уровня экспрессии гена дикого типа (дополнительная таблица S2, дополнительные материалы онлайн).Мы также проанализировали расширенный набор нарушений окружающей среды, и результаты были аналогичными (дополнительная таблица S3, дополнительные материалы онлайн). После удаления сильно коррелированных признаков экспрессии (см. «Материалы и методы») мы по-прежнему наблюдали качественно аналогичные результаты для 54 оставшихся признаков (дополнительная таблица S4, дополнительные материалы онлайн).
Таблица 1. Ранговая корреляцияСпирмена между важностью признака экспрессии гена для приспособленности и средним размером эффекта делеции гена (| ES G |) или возмущения окружающей среды (| ES E |).
| Коррелированные переменные . | Управляемые переменные . | Спирмена ρ . | P Значение . |
|---|---|---|---|
| Фитнес-эффект a , | ES E | | −0,146 | 2,1e − 16 | |
| Фитнес-эффект a , | ES G | | −0.180 | 3.7e − 24 | |
| Фитнес-эффект a , | ES E | | | ES G | | −0,094 | 1.3e − 07 |
| Фитнес-эффект a , | ES G | | | ES E | | −0,141 | 1.9e − 15 |
| Сущность b , | ES E | | −0,088 | 8.1e − 07 | |
| Сущность b , | ES G | | −0.125 | 2.2e − 12 | |
| Сущность b , | ES E | | | ES G | | −0,051 | 4.8e − 03 |
| Сущность b , | ES G | | | ES E | | −0,103 | 9,6e − 09 |
| Коррелированные переменные . | Управляемые переменные . | Спирмена ρ . | P Значение . |
|---|---|---|---|
| Фитнес-эффект a , | ES E | | −0,146 | 2,1e − 16 | |
| Фитнес-эффект a , | ES G | | −0,180 | 3,7e − 24 | |
| Фитнес-эффект a , | ES E | | | ES G | | −0,094 | 1.3e − 07 |
| Фитнес-эффект a , | ES G | | | ES E | | -0.141 | 1.9e − 15 |
| Сущность b , | ES E | | −0,088 | 8.1e − 07 | |
| Сущность b , | ES G | | −0,125 | 2.2e − 12 | |
| Сущность b , | ES E | | | ES G | | −0,051 | 4.8e − 03 |
| Сущность b , | ES G | | | ES E | | -0.103 | 9.6e − 09 |
Ранговая корреляция Спирмена между важностью признака экспрессии гена для приспособленности и средним размером эффекта делеции гена (| ES G |) или возмущения окружающей среды (| ES E |).
| Коррелированные переменные . | Управляемые переменные . | Спирмена ρ . | P Значение . |
|---|---|---|---|
| Фитнес-эффект a , | ES E | | −0,146 | 2,1e − 16 | |
| Фитнес-эффект a , | ES G | | −0,180 | 3,7e − 24 | |
| Фитнес-эффект a , | ES E | | | ES G | | −0,094 | 1.3e − 07 |
| Фитнес-эффект a , | ES G | | | ES E | | -0.141 | 1.9e − 15 |
| Сущность b , | ES E | | −0,088 | 8.1e − 07 | |
| Сущность b , | ES G | | −0,125 | 2.2e − 12 | |
| Сущность b , | ES E | | | ES G | | −0,051 | 4.8e − 03 |
| Сущность b , | ES G | | | ES E | | -0.103 | 9,6e − 09 |
| Коррелированные переменные . | Управляемые переменные . | Спирмена ρ . | P Значение . |
|---|---|---|---|
| Фитнес-эффект a , | ES E | | −0,146 | 2,1e − 16 | |
| Фитнес-эффект a , | ES G | | −0.180 | 3.7e − 24 | |
| Фитнес-эффект a , | ES E | | | ES G | | −0,094 | 1.3e − 07 |
| Фитнес-эффект a , | ES G | | | ES E | | −0,141 | 1.9e − 15 |
| Сущность b , | ES E | | −0,088 | 8.1e − 07 | |
| Сущность b , | ES G | | −0.125 | 2.2e − 12 | |
| Сущность b , | ES E | | | ES G | | −0,051 | 4.8e − 03 |
| Сущность b , | ES G | | | ES E | | -0,103 | 9,6e-09 |
Поскольку основные гены, как правило, не имеют канонического ТАТА-бокса в своих промоторах (Хан и др., 2013) и поскольку уровни экспрессии генов без ТАТА менее шумны, менее чувствительны к изменениям окружающей среды и более консервативны среди видов, чем гены, содержащие ТАТА (Newman et al.2006; Тирош и др. 2006), возникает вопрос, являются ли вышеуказанные результаты артефактами, вызванными ковариацией как существенности генов, так и нечувствительности к экспрессии при отсутствии TATA-боксов. Другими словами, мы можем наблюдать отрицательную корреляцию между TI и | ES E | и / или между TI и | ES G | если определенные гены должны использовать промоторы без TATA по причинам, отличным от экологической / генетической устойчивости. Чтобы исключить эту возможность, мы проанализировали гены, содержащие ТАТА, и гены без ТАТА (Rhee, Pugh, 2012) отдельно.Мы обнаружили, что гипотеза об адаптивном происхождении генетической устойчивости поддерживается как для генов, содержащих ТАТА, так и для генов без ТАТА (дополнительная таблица S5, дополнительные материалы онлайн). Доказательства независимого адаптивного происхождения устойчивости к окружающей среде ослаблены для генов, содержащих TATA, но остаются убедительными для генов без TATA (дополнительная таблица S5, дополнительные материалы онлайн). Взятые вместе, эти анализы подтверждают, что сигналы об адаптивной генетической и экологической устойчивости признаков экспрессии генов являются подлинными.
Обсуждение
Используя общегеномную обратную генетику, мы оценили долю несущественных генов, впервые влияющих на признак для большого количества признаков в любом организме. Мы обнаружили, что эта фракция составляет в среднем 6% для 220 исследованных морфологических признаков дрожжей. Анализ 3116 признаков экспрессии дрожжевых генов показал сопоставимую степень генетической сложности. Интересно отметить, что доля генов, влияющих на признак, одинакова у дрожжей, мух и мышей ( P > 0.05 во всех попарных сравнениях; Тест Манна – Уитни U ; инжир. 1), несмотря на довольно небольшой объем данных по последним двум видам и множественные различия в фенотипировании, размере выборки и типе исследуемых признаков. Заманчиво предположить, что наши наблюдения на дрожжах, одноклеточном эукариоте, могут быть широко применимы к другим организмам, включая млекопитающих. Однако необходимы дополнительные исследования, чтобы подтвердить это наблюдение.
Хотя доля генов, влияющих на признак дрожжей, в среднем кажется промежуточной (6% или ~ 300 несущественных генов), вариация этого количества между признаками огромна.Почти две пятых признаков относительно просты, на каждый из них влияет <1% генов (т.е. ~ 50 генов). Две пятых признаков имеют средний уровень сложности, на каждый из них влияет 1–10% генов (т. Е. 50–500 генов). Более одной пятой признаков являются очень сложными, на каждый из них влияет> 10% генов (т. Е.> 500 генов), в том числе те, на которые влияют> 30% генов (т. Е.> 1500 генов). Системный подход (Mackay et al. 2009) не только предпочтителен, но и необходим для понимания того, почему и как такое количество генов влияет на каждую из этих очень сложных черт.Теоретические исследования необходимы, чтобы понять, как обнаруженное распределение генетической сложности фенотипических признаков влияет на фенотипические вариации и эволюцию.
Наши результаты частично объясняют, почему прямая генетика неэффективна при картировании генотип-фенотип. Среди большого количества генов, которые потенциально могут влиять на сложный признак, обычно только несколько являются вариабельными в популяции картирования каждого анализа сцепления. В ассоциативных исследованиях, хотя количество вариабельных причинных генов может быть большим, когда картируемая популяция большая, статистическая мощность, как правило, низкая, потому что большинство причинных генов не очень полиморфны.Если кто-то интересуется фактическими мутациями, вызывающими ту или иную вариацию признака в популяции, использование прямой генетики кажется необходимым. Но если кто-то интересуется молекулярно-генетической сетью, ответственной за признак и потенциально влияющей на него, обратная генетика предлагает более полный и беспристрастный взгляд. Хотя здесь рассматриваются только делеции генов, полногеномная обратная генетика применима к другим типам мутантов, когда они становятся доступными, включая мутанты с избыточной функцией.
Обычный подход к верификации причинного гена-кандидата, идентифицированного прямой генетикой, заключается в изучении фенотипического эффекта удаления гена из штамма дикого типа.Однако проверка может только доказать, что ген влияет на признак, но не может подтвердить, что ген вызывает вариацию признака, наблюдаемую в популяции картирования. Это особенно верно для очень сложных признаков, когда случайно выбранный ген имеет> 10% шанс повлиять на признак. Для уменьшения количества ложноположительных результатов потребуются дополнительные тесты, такие как замена аллелей.
Мы показали, что фенотипическая изменчивость (CV) признака среди изогенных особей дикого типа уменьшается с ростом TI, что согласуется с гипотезой естественного отбора в отношении устойчивости к окружающей среде / стохастической устойчивости.Мы также показали, что средний размер эффекта делеции гена уменьшается по мере того, как признак становится более важным, что согласуется с гипотезой естественного отбора в отношении генетической устойчивости. Мы обнаружили, что экологическая / стохастическая устойчивость и генетическая устойчивость не могут полностью объяснить друг друга, отвергая гипотезу конгруэнтности (де Виссер и др., 2003) и предполагая разное происхождение двух типов устойчивости. Одним из обоснований гипотезы конгруэнтности является то, что некоторые гены, лежащие в основе устойчивости к окружающей среде, также используются для генетической устойчивости (Lehner 2010).Часто цитируемым примером является белок теплового шока Hsp90 в Drosophila (Meiklejohn and Hartl 2002). Но более поздняя работа обнаружила, что буферизация Hsp90 генетических возмущений не зависит от экологической / стохастической устойчивости (Milton et al. 2003). Более того, данные картирования мышей, Arabidopsis и дрожжей показали, что генетическая устойчивость и устойчивость к окружающей среде часто контролируются разными локусами (Fraser and Schadt 2010). Хотя генетическая устойчивость может также происходить из некоторых внутренних свойств сетей взаимодействия генов без прямого отбора на устойчивость (Siegal and Bergman 2002; Hermisson and Wagner 2004), эта гипотеза не может объяснить, почему наблюдаемая генетическая устойчивость выше для более важных признаков.Взятые вместе, наши результаты предоставляют убедительные доказательства действия естественного отбора в формировании GPM и улучшении мутационной устойчивости относительно важных признаков у дрожжей.
Три причины могут объяснить, почему несколько более ранних исследований не нашли четких доказательств естественного отбора на генетическую устойчивость. Во-первых, ожидается, что естественный отбор на генетическую устойчивость будет слабым, если размер популяции не велик, а частота вредных мутаций не высока (Wagner et al. 1997).Предыдущие неоднозначные результаты по мухам (Stearns and Kawecki 1994; Stearns et al. 1995; Houle 1998) могут отражать более слабый отбор на генетическую устойчивость у мух, чем у дрожжей, из-за их разницы в эффективном размере популяции. В свете этого сравнения интересно спросить, существует ли потенциально генетическая устойчивость у людей, у которых эффективный размер популяции 10 4 и частота нулевых мутаций 1,5 × 10 -5 на ген на поколение (см. Материалы и методы).Мы рассчитали, что S = 2 N e ∑ ( µ i Δ s i / s i ) = 0,3∑ (Δ s i / s i ) для модификатора, буферирующего вредные мутации у людей. Предполагая, что среднее значение Δ s i / s i составляет 0,5, S будет превышать 1, если модификатор одновременно влияет на> 6 генов.Поскольку общее количество генов у человека примерно в четыре раза больше, чем у дрожжей, если f генов у человека не ниже, чем у дрожжей, возможно, что модификатор влияет на гораздо больше, чем шесть генов. Тем не менее, из этого расчета ясно, что отбор по генетической устойчивости у людей в ~ 100 раз слабее, чем у дрожжей. Во-вторых, TI оценивается количественно в нашем анализе, но не во многих предыдущих исследованиях, что делает наш анализ более мощным и объективным, чем предыдущие.Но следует отметить, что мы оценили морфологический TI по наклону корреляции между значениями признаков и значениями приспособленности среди 2779 штаммов с делецией гена. Таким образом, наши оценки могут неточно отражать причинно-следственные связи между вариацией признака и приспособленностью. Однако практически невозможно установить причинно-следственные связи между чертами и приспособленностью, потому что ни одна черта не является независимой от всех других черт, так что можно манипулировать чертой, не затрагивая все другие черты.Тот факт, что наше использование неточных оценок TI по-прежнему дает важные доказательства, подтверждающие естественный отбор на генетическую устойчивость, предполагает, что истинный сигнал еще сильнее. Другими словами, наши результаты, скорее всего, будут консервативными. Обратите внимание, что в некоторых более ранних исследованиях TI назывался или определялся по-другому. Например, некоторые исследователи использовали термин «чувствительность», определяемый как процент изменения приспособленности, связанный с изменением фенотипа на 1% или 10% (Stearns and Kawecki 1994; Stearns et al.1995; Houle 1998), а Proulx et al. (2007) измерили «важность» признака экспрессии гена по дефекту роста, вызванному делецией гена. Наконец, что, вероятно, наиболее важно, наши данные намного больше, чем те, которые использовались во всех предыдущих исследованиях, что позволяет выявить отбор по генетической устойчивости и исключить гипотезу конгруэнтности.
В нашем анализе есть три оговорки. Во-первых, морфологические вариации дрожжевых клеток дикого типа были измерены в одной и той же общей среде, что может недооценивать CV, что, в свою очередь, может привести к переоценке генетической устойчивости, необъяснимой устойчивостью окружающей среды.Но эта критика не относится к анализируемым здесь данным по экспрессии генов, потому что они включают грубые изменения окружающей среды. Хотя эти среды не похожи на исторические природные среды дрожжей, они включают важные переменные среды, с которыми дрожжи сталкиваются в природе, такие как температура, осмотическое давление и концентрации аминокислот. Общие сходные результаты генетической устойчивости между морфологическими и экспрессионными признаками предполагают, что отсутствие общих изменений окружающей среды оказывает минимальное влияние на результат морфологических признаков, но этот вывод требует дальнейшего подтверждения.
Во-вторых, наши измерения величины эффекта ограничиваются нулевыми мутациями, в то время как в природе также существует множество мутаций, которые влияют на функцию гена лишь незначительно или умеренно; размер их эффекта был бы меньше. Если влияние генной мутации на различные признаки пропорционально меньше, когда мутация уменьшает, но не отменяет функцию гена, все наши эмпирические результаты должны оставаться в силе. Наш расчет преимущества генетической устойчивости в пригодности является консервативным, потому что учет дополнительных вредных (но не нулевых) мутаций увеличит преимущество генетической устойчивости.По очевидным причинам наш анализ ограничивается делециями ~ 80% генов дрожжей, которые не являются существенными. Хотя мы не ожидаем, что существенные гены будут вести себя качественно иначе, чем второстепенные гены, необходимы будущие исследования, чтобы подтвердить это ожидание.
В-третьих, наше исследование было сосредоточено на лабораторных штаммах дрожжей, потому что все делеционные линии были созданы на генетическом фоне лабораторного штамма. Применимы ли наши результаты к естественным штаммам дрожжей, требует будущих исследований.Недавние исследования выявили существенные геномные (Bergstrom et al. 2014) и морфологические (Yvert et al. 2013) вариации среди штаммов дрожжей. Наш анализ может быть применен к штаммам с различным генетическим фоном, когда становятся доступными линии делеции генов в этих фонах, а также их морфологические данные.
Неизвестно, какие молекулярно-генетические механизмы ответственны за наблюдаемое сокращение масштабов воздействия экологических и генетических нарушений на важные признаки.Предыдущие исследования дрожжей выявили так называемые гены-конденсаторы, которые могут нейтрализовать фенотипические вариации при нарушениях окружающей среды. Например, было обнаружено, что гены с большим влиянием на приспособленность при делеции с большей вероятностью будут конденсаторами экспрессии (Bergman and Siegal 2003), а гены с большим количеством генетических взаимодействий скорее всего будут конденсаторами морфологии (Levy and Siegal 2008). Однако в этих исследованиях не изучали, изменяется ли буферное воздействие на признак в зависимости от TI. Следовательно, роль этих конденсаторов в выявленной здесь адаптивной генетической и экологической устойчивости неясна.
Наблюдаемая мутационная устойчивость GPM — палка о двух концах. С одной стороны, это уменьшает вредное воздействие мутаций на важные признаки, так что уменьшается серьезность связанных дефектов. С другой стороны, из-за уменьшения размера эффекта дефекты менее вредны и, следовательно, имеют тенденцию более широко распространяться в популяции. Полные ответвления GPM, устойчивых к мутациям, ждут дальнейшего изучения, как и молекулярные механизмы, обеспечивающие устойчивость.
Материалы и методы
Морфологические и фитнес-данные
Фенотипические данные 501 морфологического признака, измеренного в штаммах дрожжей дикого типа (123 репликативных популяции) и 4718 штаммах дрожжей с делецией несущественных генов (каждый с одной популяцией) в богатой среде YPD (дрожжевой экстракт, пептон и декстроза) были получены Ohya. и другие. (2005). Мы сосредоточились на 220 из 501 признака, потому что эти 220 признаков были измерены в отдельных клетках, тогда как другие признаки были измерены для популяций клеток.Для нашего анализа необходимо использовать одноклеточные измерения. В дополнительном наборе данных S1, Дополнительный материал онлайн, перечислены эти 220 признаков, а также их CV, f гены , среднее | ES |, среднее чистое | ES | и значения TI. Значения пригодности YPD делеционных штаммов относительно дикого типа были недавно измерены Qian et al. (2012).
Фракция генов, влияющих на признак
Результаты на мышах (рис. 1 a ) были взяты из сводки исследований нокаута гена (Flint and Mackay 2009).Результаты по плодовой мушке (рис. 1 b ) были основаны на ранее опубликованных данных по линиям внедрения Р-элементов (Mackay 2010). Поскольку каждая строка обычно содержит несколько вставок P-элемента, мы вычислили долю вставок одного P-элемента, которая влияет на признак, используя среднее количество вставок на строку (Mackay et al. 1992) в предположении отсутствия эпистаза.
В статистическом анализе данных о дрожжах считается, что ген влияет на признак, когда штамм с делецией гена и штамм дикого типа имеют значительную разницу в среднем значении признака.Сначала мы рассчитали значение P путем сравнения нескольких клеток каждой линии делеции и клеток произвольно выбранной популяции репликатов дикого типа (04his3-1) с помощью теста Манна-Уитни U . Значение P <5% использовалось для установления статистической значимости. Таким образом, мы получили фракцию мутантов, у которых этот признак затронут ( f mt ). Для контроля ложноположительных результатов мы аналогичным образом провели тест Манна-Уитни U между 04his3-1 и каждой из других 122 популяций дикого типа и рассчитали фракцию ( f wt ) 122 популяций дикого типа. в котором признак значительно отклоняется от 04his3-1.Оценка доли генов, влияющих на признак ( f генов ), равна f mt — f wt , если f mt > f wt ; в противном случае мы устанавливаем f генов = 0.
Менее коррелированные морфологические признаки
Чтобы проверить, были ли некоторые из наших результатов вызваны сильно коррелированными признаками, мы попытались удалить генетически сильно коррелированные признаки.Мы измерили генетическую корреляцию между парой признаков, сопоставив значения их признаков по 4718 штаммам с делециями генов. Затем мы удалили черты одну за другой из тех, у которых были самые высокие абсолютные корреляции, до тех пор, пока ни у каких двух черт не было корреляции Пирсона, абсолютное значение которой больше 0,7. Окончательный набор данных имеет 54 признака, а распределение генов f показано на дополнительном рисунке S2, Дополнительный материал онлайн. Поскольку морфологические признаки в некоторой степени естественным образом коррелированы, еще предстоит определить, лучше ли исходные 220 признаков или 54 менее коррелированных признака лучше представлять случайно выбранные признаки.54 менее коррелированных признака также были использованы в дополнительном рисунке S5, Дополнительный материал в Интернете.
Raw ES и Net ES
Необработанный ES (ES ij ) удаления гена i по признаку j определяется как ( x ij — w j ) / w j , где x ij — среднее фенотипическое значение признака j в делеционном штамме i , а w j — соответствующее значение для дикого типа (усредненное по всем реплицированным популяциям).Условно ES ij определяется как ( x ij — w j ) / SD j , где SD j — стандартное отклонение признака в дикого типа (Mackay et al. 2009). Мы избегали использования традиционного определения, потому что ожидаемое значение SD j в значительной степени определяется точностью измерения признака, что позволяет сравнить среднее значение | ES | среди признаков в первую очередь сравнение качества измерения, а не биологии признаков.Напротив, ожидаемое значение w j не зависит от неточности измерения.
Оценить нетто | ES | удаления гена признака, мы создали 1000 наборов псевдофенотипических данных. Чтобы сгенерировать набор псевдоданных, мы случайным образом выбрали одну реплицитную популяцию дикого типа и выбрали (с заменой) из этой популяции то же количество клеток, что и в фактических данных об удалении гена. Затем мы вычислили псевдо | ES | для каждого из этих наборов псевдоданных и вычислили его среднее значение.Поскольку было сгенерировано 1000 наборов псевдоданных, фактически были использованы все 123 популяции дикого типа. Net | ES | равно raw | ES | минус среднее псевдо | ES | если сырые | ES | > среднее псевдо | ES |; в противном случае net | ES | = 0.
Фенотипическая вариация
Фенотипические вариации у дикого типа измеряли с помощью CV. Мы рассчитали дисперсию между клетками в каждой повторной популяции и усреднили ее по 123 популяциям ( V в пределах ). Затем мы рассчитали дисперсию между средними фенотипическими значениями 123 популяций ( V между )., где м — среднее из средних фенотипических значений для 123 популяций.
Относительный ТИ
Для признака j мы провели линейную регрессию F i = a j — b j (net | ES ij |) для всех i , где net | ES ij | представляет собой абсолютное значение чистой ES делеции гена i по признаку j и F i является YPD-приспособленностью штамма, лишенного гена i , по сравнению с диким типом (Qian et al.2012). Перехват a j является ожидаемой пригодностью, когда net | ES ij | = 0, тогда как наклон b j > 0 — это в 100 раз снижение приспособленности, вызванное 1% изменением фенотипической ценности признака j . Таким образом, b j является мерой относительной важности признака j для приспособленности, или TI. Поскольку в этой модели мы сосредоточились на вредных мутациях, мы использовали только те гены, которые при удалении снижают приспособляемость.Мы также попробовали журнал модели log F i = a j — b j (net | ES ij |) и обнаружили, что результаты аналогичны (дополнительный рис. S7 , Дополнительные материалы на сайте).
В приведенной выше оценке TI мы предположили, что для признака положительный эффект и отрицательный эффект одинакового размера имеют одинаковый эффект приспособленности, что может не быть верным для всех признаков. Поскольку положительные и отрицательные эффекты определяются произвольно, мы также учитывали только положительные (или только отрицательные) исходные эффекты в последующем анализе (дополнительная таблица S1, дополнительные материалы онлайн).
Анализ главных компонентов сети |
ES | МатрицаЧтобы проверить, влияет ли отсутствие зависимости между чертами на наши результаты, мы, следуя предыдущему исследованию (Wang et al. 2010), провели анализ главных компонентов для преобразования сети | ES | матрица M (4718 генов × 220 признаков). Анализ главных компонент был выполнен функцией «princomp» в MATLAB. После того, как эта функция вернула матрицу коэффициентов C (220 × 220), мы вычислили M ′ = MC (4718 преобразованных эффектов × 220 основных черт), что дает размер чистого эффекта каждого гена для каждого из 220 ортогональных признаков главных компонентов.Затем мы использовали M ′ для оценки TI.
Прогнозирование пригодности мутантного штамма с учетом GPM
Мы построили многомерную линейную модель приспособленности дрожжей, которая включает все 220 признаков и 2 779 штаммов с делециями генов, которые менее подходят, чем дикий тип: , где α — константа. Мы оценили α и β j для всех 220 признаков, используя функцию «glmfit» в MATLAB. Основываясь на приведенной выше формуле и оценочных значениях α и β j , мы предсказали F i при делеции гена i , когда либо исходная, либо случайно перемешанная сеть | ES ij | были использованы значения.
Скорость нулевых мутаций на ген на поколение
На основании геномных последовательностей штаммов дрожжей с накоплением мутаций (Lynch et al. 2008), частота точечных мутаций у дрожжей составляет 3,3 × 10 — 10 на сайт на поколение; частота небольших (1-3 п.н.) индел-мутаций составляет 2 × 10 −11 на сайт на поколение; а частота мутаций потери гена составляет 2,1 × 10 – 6 на ген на поколение. Взятые вместе, мы оценили частоту нулевых мутаций в гене на поколение примерно как [3.3 × 10 — 10 × (3/63) + 2 × 10 — 11 × 0,83] × 1,419 + 2,1 × 10 — 6 = 2,146 × 10 — 6 . Здесь 3/63 — это средняя вероятность того, что случайная точечная мутация в кодирующей области не имеет смысла, 0,83 — это доля небольших инделей, которые не кратны трем нуклеотидам (Zhang and Webb 2003), а 1419 — среднее количество кодирующих нуклеотидов на ген, кодирующий дрожжевой белок (Zhang 2000).
У человека частота точечной мутации равна 1.25 × 10 — 9 на сайт в год, а частота инделляционных мутаций составляет 1 × 10 — 10 на сайт в год (Zhang and Webb 2003). Основываясь на вариациях числа копий у людей, было подсчитано, что скорость потери гена составляет ∼10 — 5 на ген на поколение (Zhang et al. 2009). Таким образом, общая частота нулевых мутаций на ген на поколение составляет [1,25 × 10 — 9 × (3/63) + 1 × 10 — 10 × 0,83] × 1,341 × 25 + 1 × 10 . — 5 = 1.48 × 10 — 5 . Здесь 1341 — это среднее количество кодирующих нуклеотидов на один ген, кодирующий белок человека (Zhang 2000), а 25 — приблизительное количество лет на одно человеческое поколение.
Фитнес-преимущества модификаторов устойчивости
В диплоидной популяции, пусть A будет аллелем дикого типа в гене i и a представляют все нулевые аллели, которые предполагается полностью рецессивными для A . Значения пригодности для AA , Aa и aa равны 1, 1 и 1 — s i , соответственно, где s i > 0.Пусть частота мутаций от A до a будет μ i , а частота обратной мутации будет равна 0. При балансе мутации-отбора (Hartl and Clark 1997) равновесная частота а.о. особей равна . μ i / s i и ожидаемая приспособленность случайно выбранной особи в популяции составляет 1 × (1 — μ i / s i ) + (1 — s i ) × ( μ i / s i ) = 1 — μ i . Давайте рассмотрим модификатор устойчивости, который маскирует вредное воздействие a , так что aa человек теперь имеют приспособленность 1 — s i + Δ s i (0 <Δ s i < s i ). Ожидаемая приспособленность человека с модификатором составляет 1 × (1 — μ i / s i ) + (1 — s i + Δ s i ) × ( μ i / s i ) = 1 — μ i + μ i Δ s i / s i .Таким образом, среднее преимущество модификатора устойчивости составляет g = μ i Δ s i / s i . Если ген i влияет на несколько признаков, Δ s i / s i определяется частичным изменением его комбинированного воздействия приспособленности на эти признаки. Поскольку ожидается, что уменьшение генного эффекта на признак с большим вкладом в приспособленность будет вносить больший вклад в Δ s i , чем уменьшение такого же эффекта на признак с небольшим вкладом в приспособленность, модификаторы, которые предпочтительно уменьшают мутационные эффекты на важные признаки более выгодны, чем те, у которых нет таких предпочтений.В результате ожидается, что естественный отбор преимущественно повысит мутационную устойчивость важных признаков. Если модификатор буферизует нулевые мутации нескольких генов, его преимущество пригодности составляет ( μ i Δ s i / s i ), где ∑ указывает на суммирование по нескольким буферизованным генам. Вышеупомянутая формула также работает для вредных мутаций в целом, когда u i — это общая частота вредных мутаций в гене i , при условии, что все рассматриваемые мутации полностью рецессивны по отношению к аллелю дикого типа.
Когда a не полностью рецессивен к A , пригодность составляет 1, 1 — h i s i и 1 — s i для AA , Aa и aa , соответственно, где 0 < h <1 - преобладание a по сравнению с A . При балансе мутации и отбора (Hartl and Clark 1997) равновесная частота аллеля a составляет μ i / ( s i h i ) и ожидаемая пригодность случайного Выбранная особь в популяции составляет примерно 1-2 μ i . Давайте рассмотрим модификатор устойчивости, который маскирует вредное воздействие a , так что Aa индивидуумов теперь имеют приспособленность 1 — h i s i + h i Δ s i (0 <Δ s i < s i ) и aa индивидуумы имеют фитнес 1 — s i + Δ s i . Ожидаемая приспособленность человека с модификатором будет 1-2 μ i + 2 μ i Δ s i / s i .Таким образом, среднее преимущество модификатора устойчивости составляет 2 μ i Δ s i / s i . Если модификатор буферизует вредные мутации нескольких генов, его преимущество пригодности составляет 2∑ ( μ i Δ s i / s i ), где ∑ означает суммирование по множеству буферизованных генов.
Данные и анализ экспрессии генов
Данные по полногеномной экспрессии генов дрожжевых линий с делециями одного гена были собраны из четырех исследований (Hughes et al.2000; Hu et al. 2007; van Wageningen et al. 2010; Lenstra et al. 2011). В каждом исследовании штаммы дикого типа и штаммы с делецией гена выращивали в среде YPD или синтетической полной (SC) среде. Уровень экспрессии на микроматрице гена j в штамме, лишенном гена i , сравнивали с таковым в диком типе в той же среде, чтобы измерить влияние делеции гена i на уровень экспрессии гена j . Штаммы, лишенные более чем одного гена, не рассматривались.Если линия делеции анализировалась в нескольких исследованиях, мы использовали данные самого последнего исследования. В конце концов, данные содержали изменения экспрессии 4399 генов в 754 линиях делеций генов (дополнительный набор данных S2, дополнительные материалы онлайн). Затем мы ограничили наш анализ уровнями экспрессии 3116 (из 4399) генов, потому что эти гены снижают приспособленность при удалении (то есть приспособленность ≤ 1 независимо от статистической значимости) (Qian et al.2012).
Влияние удаления гена i на уровень экспрессии гена j определялось как ( x ij — w j ) / w j = x ij / w j — 1, где x ij — уровень экспрессии гена j в штамме без гена i и w j — уровень экспрессии гена j у дикого типа.Используемое значение x ij / w j было доступно из каждого набора данных. Поскольку данные экспрессии были получены для популяций клеток, а не для отдельных клеток, net | ES | невозможно оценить. Ошибка измерения уровня экспрессии гена на микрочипе в основном определяется уровнем экспрессии гена. Таким образом, контроль уровня экспрессии (дополнительные таблицы S2 – S5, дополнительные материалы онлайн) в значительной степени контролирует ошибку измерения.Для этих анализов использовали уровни экспрессии штамма дикого типа в YPD (Nagalakshmi et al. 2008). Мы количественно оценили влияние 35 сильно различающихся изменений окружающей среды на уровни экспрессии генов дикого типа (Proulx et al.2007), используя ту же формулу, где x ij / w j — кратное изменение экспрессия гена j индуцирована изменением окружающей среды i th. Эти 35 сред были ранее выбраны из 162 сред, представляющих наименее коррелированные экологические проблемы (Proulx et al.2007). Мы выполнили аналогичный анализ с использованием всех 162 экологических проблем, при которых ранее были измерены изменения экспрессии генов дикого типа (Gasch et al. 2000) (дополнительная таблица S3, дополнительные материалы онлайн).
При изучении потенциального воздействия сильно коррелированных признаков мы измерили генетическую корреляцию между парой признаков экспрессии, сопоставив уровни их экспрессии между мутантными штаммами. Затем мы удалили признаки экспрессии один за другим из тех, которые имели самую высокую корреляцию, до тех пор, пока ни один из двух признаков не имел корреляцию Пирсона больше 0.7. Окончательный набор данных включал 2223 признака экспрессии (дополнительная таблица S4, дополнительные материалы онлайн).
Благодарности
Мы благодарим доктора Yoshikazu Ohya за предоставленные морфологические данные о дрожжах, а также доктора Calum Maclean и Dr Jian-Rong Yang за ценные комментарии. Эта работа была частично поддержана исследовательским грантом R01GM103232, предоставленным Национальным институтом здравоохранения США J.Z.
Список литературы
. ,Введение в системную биологию: принципы проектирования биологических схем
,2007
Лондон
Chapman & Hall
,.Эволюция организации хромосом, вызванная отбором на снижение шума экспрессии генов
,Nat Genet.
,2007
, т.39
(стр.945
—949
),.Эволюционная емкость как общая характеристика сложных генных сетей
,Nature
,2003
, vol.424
(стр.549
—552
),,,,,,,,, и др.Вид в высоком разрешении функциональной генетической изменчивости из геномов природных дрожжей
,Mol Biol Evol.
,2014
, т.31
(стр.872
—888
),,.Инновации и надежность в сложных регуляторных генных сетях
,Proc Natl Acad Sci U S. A.
,2007
, vol.104
(стр.13591
—13596
),,,,,,,,, и др.Перспектива: эволюция и определение генетической устойчивости
,Evolution
,2003
, vol.57
(стр.1959
—1972
),,,.Мутационная устойчивость может способствовать адаптации
,Nature
,2010
, vol.463
(стр.353
—355
),,,,,,,,.Рассечение генетически сложных признаков с чрезвычайно большими пулами дрожжевых сегрегантов
,Nature
,2010
, vol.464
(стр.1039
—1042
),. ,Введение в количественную генетику
,1996
Лондон
Пирсон
.Эволюционная генетика канализации
,Q Rev Biol.
,2005
, т.80
(стр.287
—316
),.Генетическая архитектура количественных признаков у мышей, мух и людей
,Genome Res.
,2009
, т.19
(стр.723
—733
),.Количественная генетика фенотипической устойчивости
,PLOS One
,2010
, vol.5
стр.e8635
,,,,,,,.Программы геномной экспрессии в ответ дрожжевых клеток на изменения окружающей среды
,Mol Biol Cell.
,2000
, т.11
(стр.4241
—4257
),.Канализация в эволюционной генетике: стабилизирующая теория?
,BioEssays
,2000
, т.22
(стр.372
—380
),,,,.Полногеномная характеристика взаимосвязи между эссенциальными и TATA-содержащими генами
,FEBS Lett.
,2013
, т.587
(стр.444
—451
),. ,Принципы популяционной генетики
,1997
Сандерленд (Массачусетс)
Sinauer Associates
,.Популяционно-генетическая теория скрытой изменчивости и генетической устойчивости
,Genetics
,2004
, vol.168
(стр.2271
—2284
).Как мы должны объяснить вариацию генетической изменчивости признаков?
,Genetica
,1998
, т.102-103
(стр.241
—253
),,.Генетическая реконструкция функциональной регуляторной сети транскрипции
,Nat Genet.
,2007
, т.39
(стр.683
—687
),,,,,,,,, и др.Функциональное открытие через сборник профилей экспрессии
,Cell
,2000
, vol.102
(стр.109
—126
).Селекция для минимизации шума в живых системах и его значение для эволюции экспрессии генов
,Mol Syst Biol.
,2008
, т.4
стр.170
.Гены придают аналогичную устойчивость к средовым, стохастическим и генетическим возмущениям у дрожжей
,PLOS One
,2010
, vol.5
стр.e9035
,,,,,,,,, и др.Специфичность и топология путей взаимодействия хроматина в дрожжах
,Mol Cell.
,2011
, т.42
(стр.536
—549
),.Сетевые концентраторы буферизуют изменения окружающей среды в Saccharomyces cerevisiae
,PLOS Biol.
,2008
, т.6
(стр.2588
—2604
),,,.Влияние вставок одиночных Р-элементов на количество щетинок и жизнеспособность у Drosophila melanogaster
,Genetics
,1996
, vol.143
(стр.277
—292
),,,,,,,,, и др.Полногеномный вид спектра спонтанных мутаций у дрожжей
,Proc Natl Acad Sci U S A.
,2008
, т.105
(стр.9272
—9277
).Мутации и количественная генетическая изменчивость: уроки Drosophila
,Phil Trans R Soc B.
,2010
, vol.365
(стр.1229
—1239
),,.Влияние вставок Р-элемента на количественные признаки у Drosophila melanogaster
,Genetics
,1992
, vol.130
(стр.315
—332
),,.Генетика количественных признаков: проблемы и перспективы
,Nat Rev Genet.
,2009
, т.10
(стр.565
—577
).Генетическая архитектура количественных признаков
,Annu Rev Genet.
,2001
, т.35
(стр.303
—339
),,,,,,,,, и др.Обнаружение недостающей наследственности сложных болезней
,Nature
,2009
, vol.461
(стр.747
—753
).Вариация и выбор полигенных признаков
,J Genet.
,1941
, т.41
(стр.159
—193
),.Одноместный режим канализации
,Trends Ecol Evol.
,2002
, т.17
(стр.468
—473
),,,,.Количественная симметрия признака, не зависящая от буферизации Hsp90: различные способы генетической канализации и стабильности развития
,Proc Natl Acad Sci U S A.
,2003
, т.100
(стр.13396
—13401
),,,,,,.Транскрипционный ландшафт генома дрожжей, определенный с помощью секвенирования РНК
,Science
,2008
, vol.320
(стр.1344
—1349
),,,,,,.Одноклеточный протеомный анализ S. cerevisiae показывает архитектуру биологического шума
,Nature
,2006
, vol.441
(стр.840
—846
),,,,,,,,, и др.Высокомасштабное и крупномасштабное фенотипирование дрожжевых мутантов
,Proc Natl Acad Sci U S. A.
,2005
, vol.102
(стр.19015
—19020
),,.Прямой отбор по генетической устойчивости, выявленной в дрожжевом транскриптоме
,PLOS One
,2007
, vol.2
стр.e911
,,,,.Геномный ландшафт и эволюционное разрешение антагонистической плейотропии у дрожжей
,Cell Rep.
,2012
, т.2
(стр.1399
—1410
),,,,,,,,.Большинство генов животных необходимы для приспособленности к дикому типу
,Cell
,2012
, vol.148
(стр.792
—802
),.Полногеномная структура и организация преинициационных комплексов эукариот
,Nature
,2012
, vol.483
(стр.295
—301
)..Природа количественной генетической изменчивости
,Наследие от Менделя
,1967
Мэдисон (Висконсин)
The Univeristy of Wisconsin Press
,,,,.Селекция на устойчивость к мутагенизированным РНК-вирусам
,PLOS Genet.
,2007
, т.3
стр.е93
.Канализация: генетические аспекты и аспекты развития
,Annu Rev Ecol Syst.
,1991
, т.22
(стр.65
—93
),.Повторный визит к канализации Ваддингтона: стабильность развития и эволюция
,Proc Natl Acad Sci U S. A.
,2002
, vol.99
(стр.10528
—10532
),,.Дифференциальная генетическая и экологическая канализация компонентов приспособленности у Drosophila melanogaster
,J Evol Biol.
,1995
, т.8
(стр.539
—557
),.Фитнес-чувствительность и канализация жизненно важных черт
,Evolution
,1994
, vol.48
(стр.1438
—1450
),,,.Генетическая подпись межвидовых вариаций экспрессии генов
,Nat Genet.
,2006
, т.38
(стр.830
—834
),,,,,,,,, и др.Функциональное перекрытие и регуляторные связи формируют генетические взаимодействия между сигнальными путями
,Cell
,2010
, vol.143
(стр.991
—1004
).Канализация развития и наследования приобретенных признаков
,Nature
,1942
, vol.150
(стр.563
—565
).Энергетические ограничения эволюции экспрессии генов
,Mol Biol Evol.
,2005а
, т.22
(стр.1365
—1374
). ,Устойчивость и эволюционируемость живых систем
,2005b
Princeton (NJ)
Princeton University Press
,,.Популяционно-генетическая теория канализации
,Evolution
,1997
, vol.51
(стр.329
—347
),.Плейотропная структура карты генотип-фенотип: эволюционируемость сложных организмов
,Nat Rev Genet.
,2011
, т.12
(стр.204
—213
),,.Геномные паттерны плейотропии и эволюция сложности
,Proc Natl Acad Sci U S A.
,2010
, т.107
(стр.18034
—18039
),.Влияние шума экспрессии генов на приспособленность организма и эффективность естественного отбора
,Proc Natl Acad Sci U S. A.
,2011
, vol.108
(стр.E67
—E76
),,,,.Эволюция цифровых организмов при высокой частоте мутаций приводит к выживанию самых плоских
,Nature
,2001
, vol.412
(стр.331
—333
),,,,,,,,, и др.Функциональная характеристика генома S. cerevisiae с помощью делеции гена и параллельного анализа
,Science
,1999
, vol.285
(стр.901
—906
),,,,,,.Феномика единичных клеток выявляет внутривидовые вариации фенотипического шума у дрожжей
,BMC Syst Biol.
,2013
, т.7
стр.54
,,,.Изменение количества копий в состоянии здоровья, болезней и эволюции человека
,Annu Rev Genomics Hum Genet.
,2009
, т.10
(стр.451
—481
).Распределение длин белков для трех областей жизни
,Trends Genet.
,2000
, т.16
(стр.107
—109
),.Эволюционное ухудшение пути вомероназальной трансдукции феромона у катарринных приматов
,Proc Natl Acad Sci U S. A.
,2003
, т.100
(стр.8337
—8341
)Заметки автора
© Автор, 2014. Опубликовано Oxford University Press от имени Общества молекулярной биологии и эволюции. Все права защищены. Для получения разрешений обращайтесь по электронной почте: journals.permissions@oup.com
. .