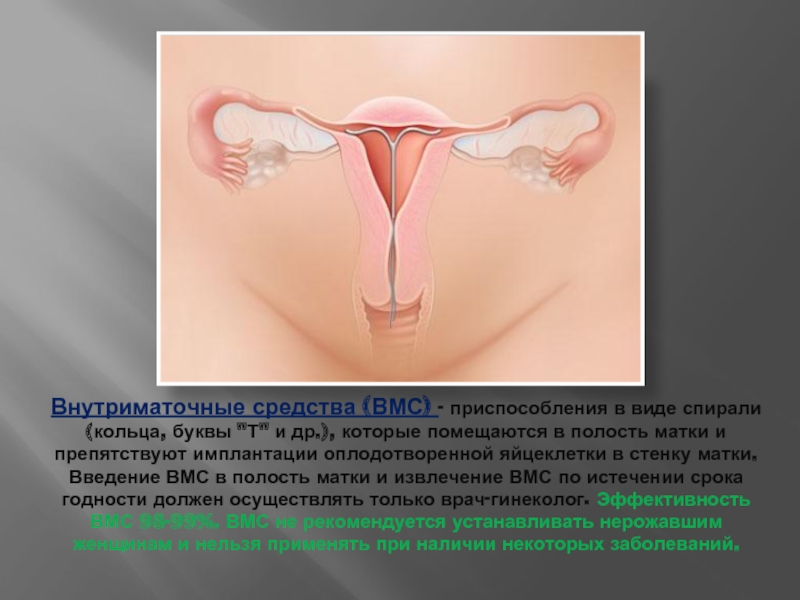
Установка ВМС в гинекологической клинике КСТ в Москве
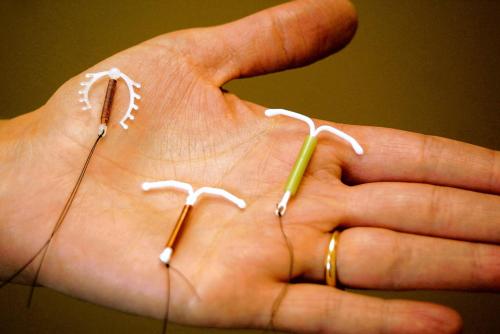
Если Вы задумались об установке внутриматочной спирали — тогда эта статья для Вас. Внутриматочная спираль — метод контрацепции, при котором большинство сперматозоидов гибнет в полости матки, а оплодотворенная яйцеклетка не прикрепляется к стенкам матки, и беременность не наступает. В зависимости от формы спирали бывают Т-образные, круглые и овальные. По составу делятся на медьсодержащие, серебросодержащие, золотосодержащие и гормональные.
Информацию о том, какую спираль выбрать, не стоит искать на женских форумах — лучше доверить этот вопрос врачу гинекологу. Контрацептивное средство подбирается индивидуально с учетом физиологических особенностей и состояния репродуктивной системы пациентки.
В нашей клинике установка внутриматочной спирали — это полноценная услуга, включающая в себя:
- консультацию;
- осмотр гинекологом;
- УЗ исследование;
- мазок на бактериальную флору;
- установку спирали;
- обучение уходу и наблюдению за контрацептивом.

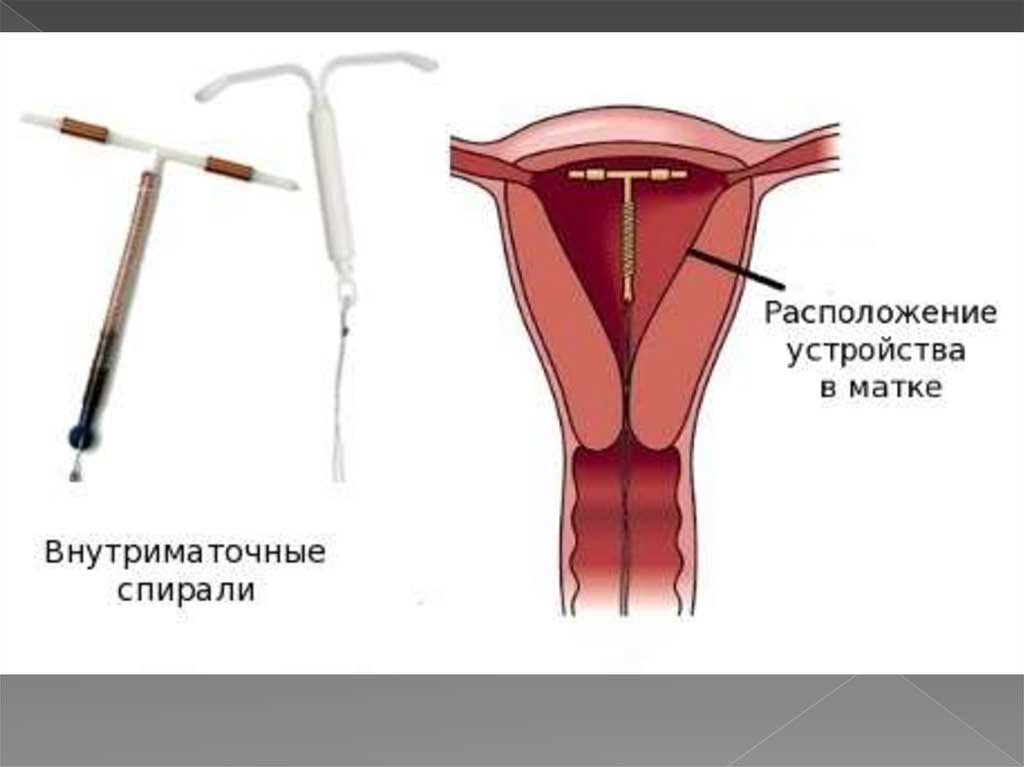
Показания для установки внутриматочной спирали
Установка спирали в матку в большинстве случаев мы рекомендуем женщинам от 35 лет, у которых есть постоянный сексуальный партнер. ВМС — альтернатива оральным контрацептивам. Таблетки меняют гормональный фон ради предотвращения беременности, их неконтролируемый, длительный прием может привести к негативным процессам как в органах малого таза так и всего организма. ВМС препятствует беременности без изменения гормонального фона, поэтому не вызывает таких осложнений.
Запись на приём
″Обратите внимание! Внутриматочная спираль повышает уязвимость половых органов перед возбудителями инфекций. Чтобы не допустить осложнений, я ввожу контрацептив в матку после обследования, сдачи анализов на мочеполовые инфекции и лечения воспалительных процессов в органах малого таза.
Врач акушер-гинеколог Шустова Ольга Леонидовна
″Обратите внимание! Внутриматочная спираль повышает уязвимость половых органов перед возбудителями инфекций. Чтобы не допустить осложнений, я ввожу контрацептив в матку после обследования, сдачи анализов на мочеполовые инфекции и лечения воспалительных процессов в органах малого таза.
Чтобы не допустить осложнений, я ввожу контрацептив в матку после обследования, сдачи анализов на мочеполовые инфекции и лечения воспалительных процессов в органах малого таза.
Врач акушер-гинеколог
Шустова Ольга Леонидовна
Запись на приём
Этот метод контрацепции подходит курящим женщинам, диабетикам, гипертоникам, женщинам с нарушением жирового обмена, высоким риском опухолей груди, матки или шейки матки.
Если хочется установить спираль…
- Поговорите с врачом, которому доверяете. Узнайте, с какими контрацептивами он работает и что советует в конкретном случае. Нельзя выбирать такие приспособления по рекламному описанию на упаковке или мнению подруг: нужен опытный гинеколог.
- Не ориентируйтесь на цену. Дорогой контрацептив – не значит «более эффективный» или «универсальный». Цена не имеет значения – только индивидуальные особенности вашего организма.
- Расскажите гинекологу о страхах. Например, вы боитесь «врастания в матку» или «исчезновения месячных». Врач объяснит, какие страхи обоснованны, а какие – нет.
- Подробно расскажите доктору о своем образе жизни. Вспомните о прерванных беременностях. Сообщите, если имеете нескольких половых партнеров, не забудьте предупредить о лечении ЗППП в прошлом. Скажите, если наблюдаетесь у эндокринолога. Врач должен иметь максимально полную картину, чтобы назначить контрацептив и гарантировать вашу безопасность на следующие годы.
 Врач объяснит, какие страхи обоснованны, а какие – нет.
Врач объяснит, какие страхи обоснованны, а какие – нет.Противопоказания к введению ВМС
Установка спирали — не для вас, если вы:
- нерожавшая женщина до 35 лет;
- имеете диагнозы «гиперплазия эндометрия» или «эрозия шейки матки»;
- страдаете аллергией на медь;
- лечили воспаления внутренних половых органов после родов;
- удалили трубу или обе по причине внематочной беременности;
- занимаетесь сексом от случая к случаю или с разными партнерами;
- лечите диабет или принимаете таблетки с глюкокортикостероидами;
- подозреваете у себя беременность;
- наблюдаете или лечите опухоль половых органов;
- замечали сукровичные выделения или полноценное кровотечение из влагалища вне менструации.

Не рекомендуется устанавливать внутриматочную спираль женщинам с болезнями крови — например, с анемией или плохой свертываемостью. ВМС также не ставят при наличии острых воспалительных процессов в организме.
Как происходит установка спирали в матку
Пациентка усаживается в гинекологическое кресло. Врач вынимает ВМС из стерильной упаковки. Затем смазывает шейку матки обезболивающим средством и ставит приспособление.
8 из 10 женщин при введении контрацептива не ощущают ничего. 2 из 10 чувствуют едва заметный короткий дискомфорт, который исчезает через несколько секунд. Некоторым пациенткам гинеколог назначает антибиотики, если видит склонность к воспалительным заболеваниям малого таза.
Процедура занимает три минуты.
В какой период цикла ставят внутриматочную спираль
Установка спирали женщине возможна в любой день менструального цикла. Из гигиенических соображений лучше не приходить на процедуру в первые дни месячных. Но оптимальный период с 4 по 8 день цикла. В это время болевые ощущения самые слабые, шейка матки приоткрыта и низкая вероятность незамеченной беременности.
В это время болевые ощущения самые слабые, шейка матки приоткрыта и низкая вероятность незамеченной беременности.
После аборта можно вставлять контрацептив сразу. После естественных родов приспособление ставят на 2-3 месяц после осмотра в гинекологическом кресле. После кесарева сечения выжидают полгода.
Подготовка к установке ВМС
- Анализы. Гинеколог направляет пациентку на анализы крови и делает мазок для изучения микрофлоры влагалища, чтобы выявить возбудителей и назначить лечение инфекций, передающихся половым путём. Еще доктор назначает тест на беременность.
- Подбор изделия. Врач задает вопросы о самочувствии, делает УЗИ полости матки и яичников, чтобы исключить опухоли и выбрать тип ВМС.
- Введение. Доктор выбирает день цикла, когда установка спирали будет наименее дискомфортной для конкретной пациентки.
″Обратите внимание! Воздержитесь от секса за 1-2 дня перед введением внутриматочного контрацептива. Не спринцуйтесь, не пользуйтесь средствами для интимной гигиены — подмывайтесь только водой.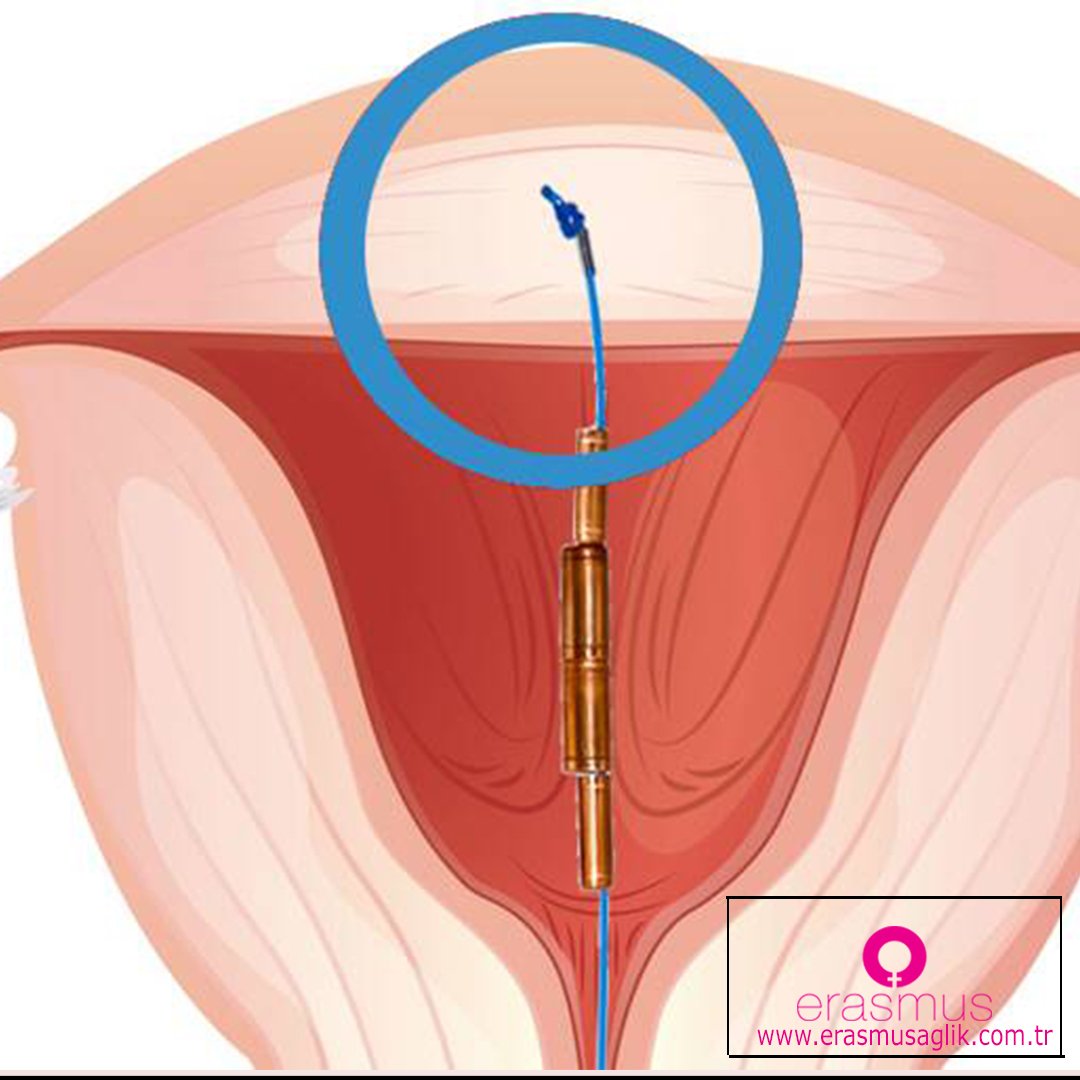
Врач акушер-гинеколог Харичкова Алефтина Михайловна
Запись на приём
″Обратите внимание! Воздержитесь от секса за 1-2 дня перед введением внутриматочного контрацептива. Не спринцуйтесь, не пользуйтесь средствами для интимной гигиены — подмывайтесь только водой. Не вставляйте вагинальные свечи и таблетки.″
Врач акушер-гинеколог Харичкова Алефтина Михайловна
Запись на приём
″Обратите внимание! Воздержитесь от секса за 1-2 дня перед введением внутриматочного контрацептива. Не спринцуйтесь, не пользуйтесь средствами для интимной гигиены — подмывайтесь только водой. Не вставляйте вагинальные свечи и таблетки.
Врач акушер-гинеколог
Харичкова Алефтина Михайловна
Запись на приём
Побочные эффекты и осложнения
Выпадение контрацептива. Иногда изделие само вываливается из матки.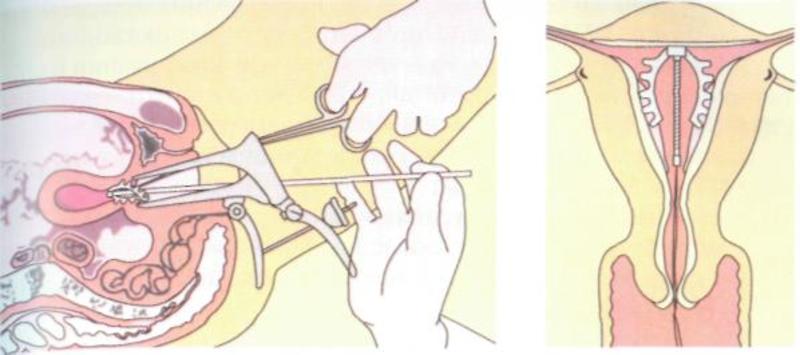 Ничего страшного: врач промахнулся с размером. Запишитесь к нему еще раз, чтобы вставить другое приспособление.
Ничего страшного: врач промахнулся с размером. Запишитесь к нему еще раз, чтобы вставить другое приспособление.
Обострение мочеполовых инфекций. С 3 по 6 месяц после процедуры могут дать знать о себе хронические заболевания внутренних половых органов. Это выражается в виде боли в пояснице или в паху, сопровождается высокой температурой и жжением во влагалище. Если такое случилось, звоните доктору. Он осмотрит вас и подберет лечение.
Маточные кровотечения и мажущие межменструальные выделения. Такое случается при воспалениях и гормональных скачках. Обратитесь к гинекологу, чтобы решить проблему.
Незначительное увеличение объема менструальной крови и усиление боли во время ПМС.
Полное исчезновение менструаций через несколько месяцев после процедуры – аменорея. Позвоните администратору клиники и обратитесь к гинекологу для лечения.
Исключительно редкие осложнения – воспаление эндометрия и прободение матки.
″Важно! ВМС не защищает от болезней, передающихся половым путем.
Врач акушер-гинеколог Харичкова Алефтина Михайловна
Запись на приём
″Важно! ВМС не защищает от болезней, передающихся половым путем. Поэтому с новым партнером дополнительно используют презерватив.″
Врач акушер-гинеколог Харичкова Алефтина Михайловна
Запись на приём
″Важно! ВМС не защищает от болезней, передающихся половым путем. Поэтому с новым партнером дополнительно используют презерватив.″
Врач акушер-гинеколог
Запись на приём
Как вынуть спираль
Самостоятельно вынимать внутриматочный контрацептив запрещено: легко повредить слизистую шейки матки и влагалища, занести инфекцию. А у доктора удалить приспособление можно в любой момент, когда нет месячных. Перед этим гинеколог проводит общий осмотр в кресле, делает мазок на флору и цитологическое исследование. Затем медицинским инструментом – корнцангом или пинцетом, вынимает спираль из матки.
Затем медицинским инструментом – корнцангом или пинцетом, вынимает спираль из матки.
Изредка для удаления требуется операция под местным наркозом.
После извлечения изделия может ныть низ живота или появиться незначительные мажущие выделения. Через пару дней выделения пройдут сами, лечения не потребуется.
Рекомендации после удаления ВМС такие же, как после установки. Возможна задержка месячных на несколько дней.
Особенности гормональных ВМС
После установки гормональных ВМС:
- Месячные могут стать короче, и крови будет меньше обычного.
- У 2 из 10 женщин менструации могут исчезнуть совсем. Это связано не со «сном» яичников, как при приеме противозачаточных таблеток, а с замедлением роста слизистой оболочки матки.
- Может пройти анемия, которая появляется при чрезмерно обильных менструациях.
- Может увеличиться вес тела. В 9 из 10 случаев это связано с питанием и работой печени. Наличие внутриматочного контрацептива влияет на массу тела косвенно.
- Уменьшится риск гиперплазии и озлокачествления опухолей эндометрия.

Перед установкой гормональных ВМС доктор направляет пациентку на анализы крови. В лаборатории проверяют взаимное соотношение половых и других гормонов. Если есть отклонения от нормы, необходимо пройти осмотр у маммолога и эндокринолога.
Особенности ухода после установки ВМС
| После установки ВМС нужно | После установки ВМС нельзя |
| Сохранять физический покой 7-10 дней. То есть не заниматься спортом и не поднимать тяжести. | Заниматься сексом – 5 дней. |
| Подмываться только мягким средством для интимной гигиены. Мыло нарушает кислотность влагалища и приводит к гибели полезной флоры. | Вставлять тампоны в первые месячные: воспользуйтесь прокладками. |
| Мыться только в душе 4-5 дней. Не принимать ванну, не ходить в баню. | Принимать Аспирин – 5 дней. |
Прийти к гинекологу на осмотр через 10 дней после процедуры. | Спринцеваться – 4 дня. |
После установки спирали желательно приходить на профилактические осмотры раз в полгода-год.
Позвоните врачу, если:
- В течение первых дней после процедуры температура поднялась до 37,2 или выше.
- Месячные задерживаются дольше, чем на 3 недели.
- Появились выделения из влагалища с неприятным запахом.
- Болит низ живота, и боль не проходит дольше 2 дней.
- Открылось маточное кровотечение.
- Изделие вывалилось.
- Менструации очень болезненные, приходится пить обезболивающие лекарства.
Частые вопросы пациентов про установку ВМС
Как часто меняют ВМС?Негормональные изделия служат 10 лет. Просто приходите на профилактический осмотр раз в полгода-год, если ничего не беспокоит. И попросите врача вытащить контрацептив, если хотите забеременеть.
Гормональные изделия меняют раз в 5 лет. Новое приспособление ставят сразу после извлечения отслужившего.
Доктор сообщит срок годности конкретного изделия, когда выберет его во время осмотра.
Можно ли забеременеть при ношении негормональной ВМС?Вероятность беременности – примерно 8 шансов из 1000 за год. 1 из 2 беременностей с ВМС заканчивается выкидышем.
Вероятность увидеть тест с двумя полосками резко увеличится, если приспособление сместится или вывалится из полости матки, а вы этого не заметите. Такое может случиться во время месячных. Поэтому проверяйте раз в месяц, на месте ли изделие. Помойте руки с мылом и нащупайте во влагалище усики. Если раньше вы находили их мигом, а теперь вдруг не нашли – сразу запишитесь к гинекологу на осмотр.
Как узнать о наступлении беременности?Беременность возможна, если месячные задерживаются дольше, чем на 2 недели. Сделайте тест и запишитесь к врачу, если он окажется положительным. Врач вынет ВМС, чтобы сохранить ребенка.
Некоторые женщины переживают, что спираль врастет в тело плода. Это миф: ребенок в матке окружен плотной оболочкой, контрацептив ему не навредит. Но лучше извлечь его, чтобы ткани матки восстановились.
Это миф: ребенок в матке окружен плотной оболочкой, контрацептив ему не навредит. Но лучше извлечь его, чтобы ткани матки восстановились.
Противозачаточный эффект от таких изделий легко обратим. В течение первого же года вероятность зачатия достигнет 96%. Месячные нормализуются в первый же цикл, ткани матки восстановятся за 2-3 месяца.
Зачатию могут воспрепятствовать мочеполовые инфекции, которые нередко обостряются при ношении спирали. Причиной бесплодия станет не контрацептив, а возбудители ЗППП – например, хламидии. Из-за этого это средство контрацепции не рекомендуют нерожавшим женщинам и тем, кто лечил бесплодие.
Когда можно планировать ребенка?На следующий месяц после извлечения контрацептива.
Если у Вас остались вопросы или Вы хотите записаться к нашему гинекологу на осмотр и консультацию, оставьте заявку на сайте или позвоните по телефону: +7 (495) 114-51-51.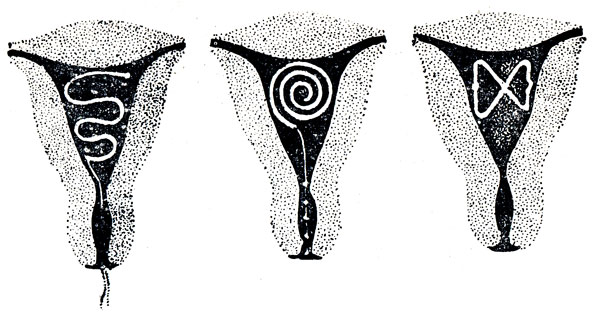 Мы перезвоним вам и подберем удобное время визита, а также напомним о записи за день до посещения клиники.
Мы перезвоним вам и подберем удобное время визита, а также напомним о записи за день до посещения клиники.
Врачи, осуществляющие установку внутриматочной спирали (ВМС)
Стоимость
| Установка внутриматочной спирали |
- Прием врача-акушера-гинеколога лечебно-диагностический, первичный, амбулаторный 1 900
- УЗИ органов малого таза женщины2 100
- Введение внутриматочной спирали3 800
- Введение ВМК «Мирена» включая стоимость контрацептива24 000
- Введение внутриматочной спирали (включая стоимость контрацептива)7 200
- Извлечение ВМС, неосложненное1 200
- Извлечение ВМС, осложненное2 800
Внутриматочная спираль — виды, как действует, как устанавливается
Внутриматочная спираль – средство контрацепции с эффективностью свыше 95%. Способ позволяет отказаться от презервативов и противозачаточных препаратов.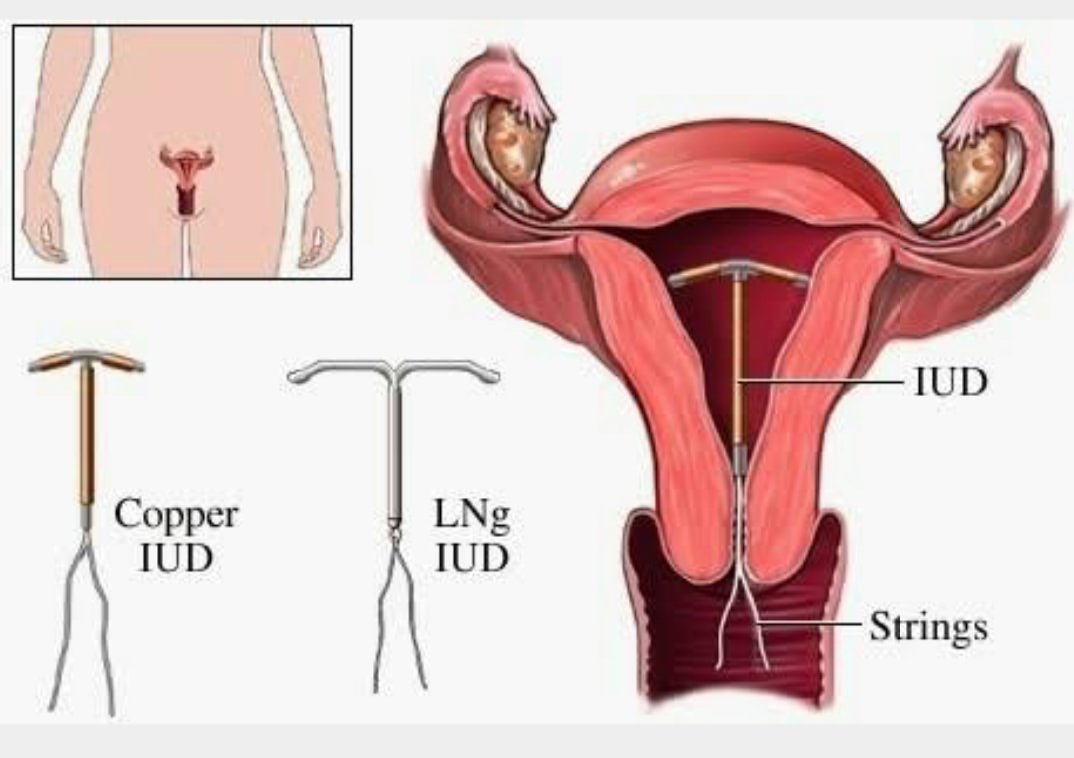
Внутриматочные спирали бывают разной формы: Т-образные, Ф-образные или в виде кольца. Чаще всего используют спирали Т-образной формы. Как правило, устройство состоит из упругих материалов: пластика и меди, поэтому легко гнется и не повреждает поверхность внутренних органов. Доступны и спирали с содержанием золота, серебра или прогестерона – гормонального компонента, блокирующего беременность. Срок действия спирали – от трех до десяти лет, в зависимости от вида и материала.
Как устанавливается
Внутриматочную спираль устанавливают, когда женщина не планирует беременность в течение ближайшего года. Перед процедурой необходимо проконсультироваться с гинекологом, чтобы выявить противопоказания и подобрать устройство. Рекомендуется устанавливать спираль с четвертого по восьмой день от начала менструального цикла. После родов – не раньше, чем через два месяца, после кесарева сечения – через полгода. Также допускается установка спирали сразу после искусственного прерывания беременности.
Установленная спираль слабо ощущается и не доставляет дискомфорт. Доктор оставляет две короткие нити из шейки матки для контроля положения устройства и будущего изъятия. Во время полового акта нити не мешают. Удалением спирали занимается врач-гинеколог. Самостоятельные попытки извлечь устройство приводят к повреждениям внутренних органов.
В течение недели после процедуры необходимо избегать физических нагрузок, воздержаться от интимной близости, горячей ванны и бани. О персональных рекомендациях уточняйте у лечащего врача.
Как действует
Спираль с медными составляющими мешает сперматозоидам проникать внутрь матки для оплодотворения яйцеклетки. Медь снижает активность сперматозоидов и предотвращает воспаления.
Спираль с гормональным компонентом производят на основе прогестерона – левоноргестрела. Гормон не дает сперматозоидам попасть в маточную полость и часто подавляет овуляцию.
Показания для установки спирали:
- рекомендация врача после родов, аборта, выкидыша;
- женщина хочет отказаться от приема гормональных препаратов;
- противозачаточные средства не работают;
- возможная беременность несет опасность для здоровья женщины.
Противопоказания для установки спирали
- Беременность.
- Внематочная беременность.
- Воспаления органов малого таза, болезни шейки матки.
- Новообразования шейки матки.
- Деформации и аномалии развития матки.
- Патологии менструального кровотечения.
- ВИЧ-инфекция.
- Венерические болезни.
- Болезни крови.
- Аллергия на материал спирали.
- Туберкулез половых органов.
- Хронические заболевания органов малого таза.
Преимущества метода
- Степень защиты от беременности – от 95%.
- Способ подходит для экстренной контрацепции после полового акта.
- Действие начинается сразу после установки.
- Долгосрочный эффект – до 10 лет.
- Не мешает при половом акте.
- Не влияет на грудное вскармливание.
Недостатки метода
- Не всегда подходит молодым и нерожавшим девушкам;
- Не предоставляет защиту от болезней, передающихся половым путем;
- Возможны изменения менструального цикла.

Возврат к списку
Установка внутриматочной спирали в СПб
Внутриматочная спираль — современный и надежный метод контрацепции, эффективность которого составляет не менее 98%. В зависимости от типа спирали контрацептивный эффект сохраняется от 3 до 10 лет.
Основной принцип действия — торможение сперматозоидов, направляющихся в полость матки. Спираль сокращает длительность жизни яйцеклетки, и противостоит прикреплению уже оплодотворенной яйцеклетки к стенке маточной трубы.
Внутриматочную спираль может установить и извлечь только врач-гинеколог.
Внутриматочные контрацептивы бывают двух видов: медьсодержащие и гормональные.
Медьсодержащие спирали имеют Т-образную форму, ее стержень и «усики» обмотаны медной проволокой. Введенное в полость матки инородное тело (спираль) не позволяет полости матки смыкаться. Кроме того, медь, вызывает небольшую местную воспалительную реакцию, в результате чего сперма теряет способность к оплодотворению.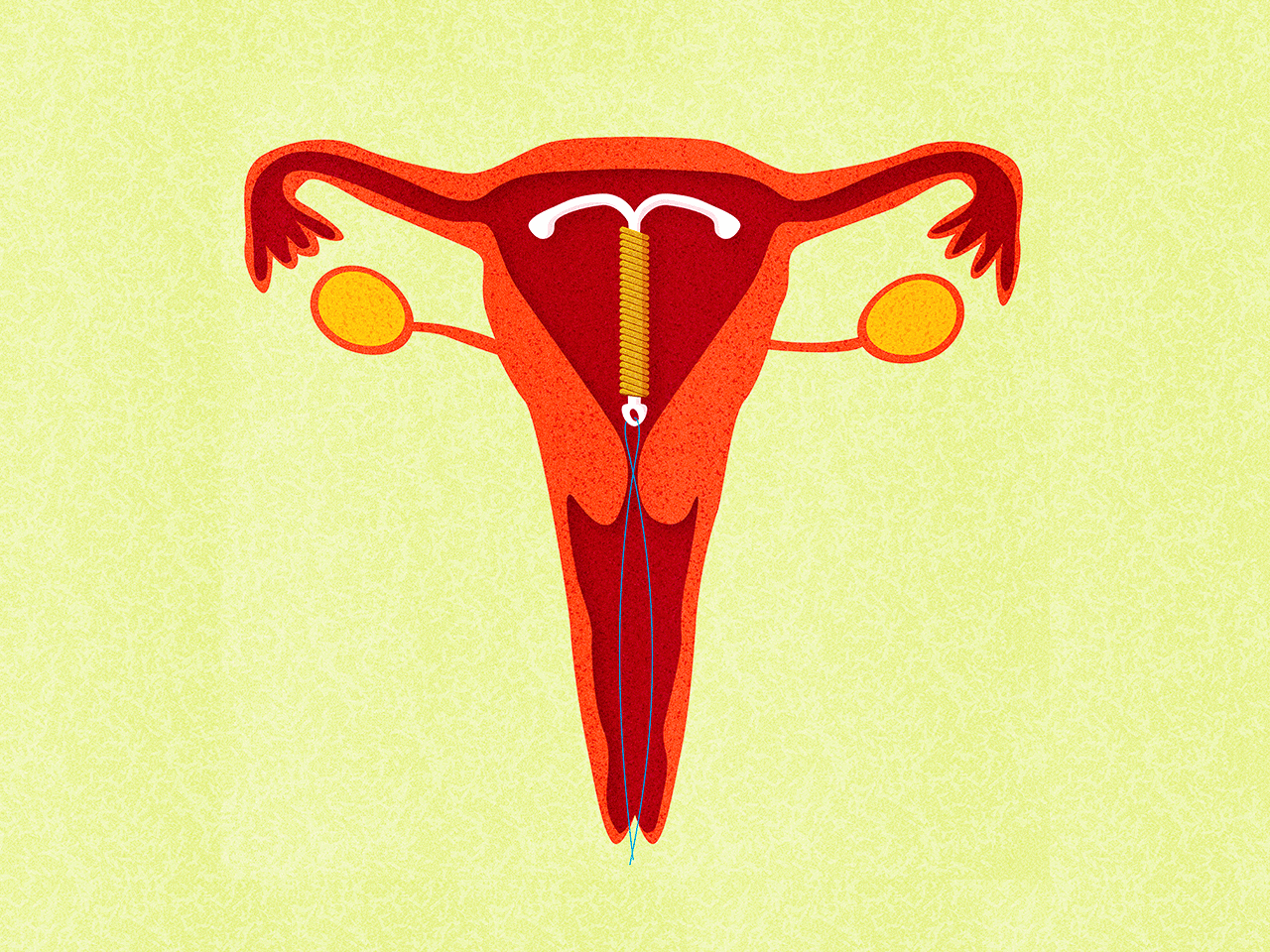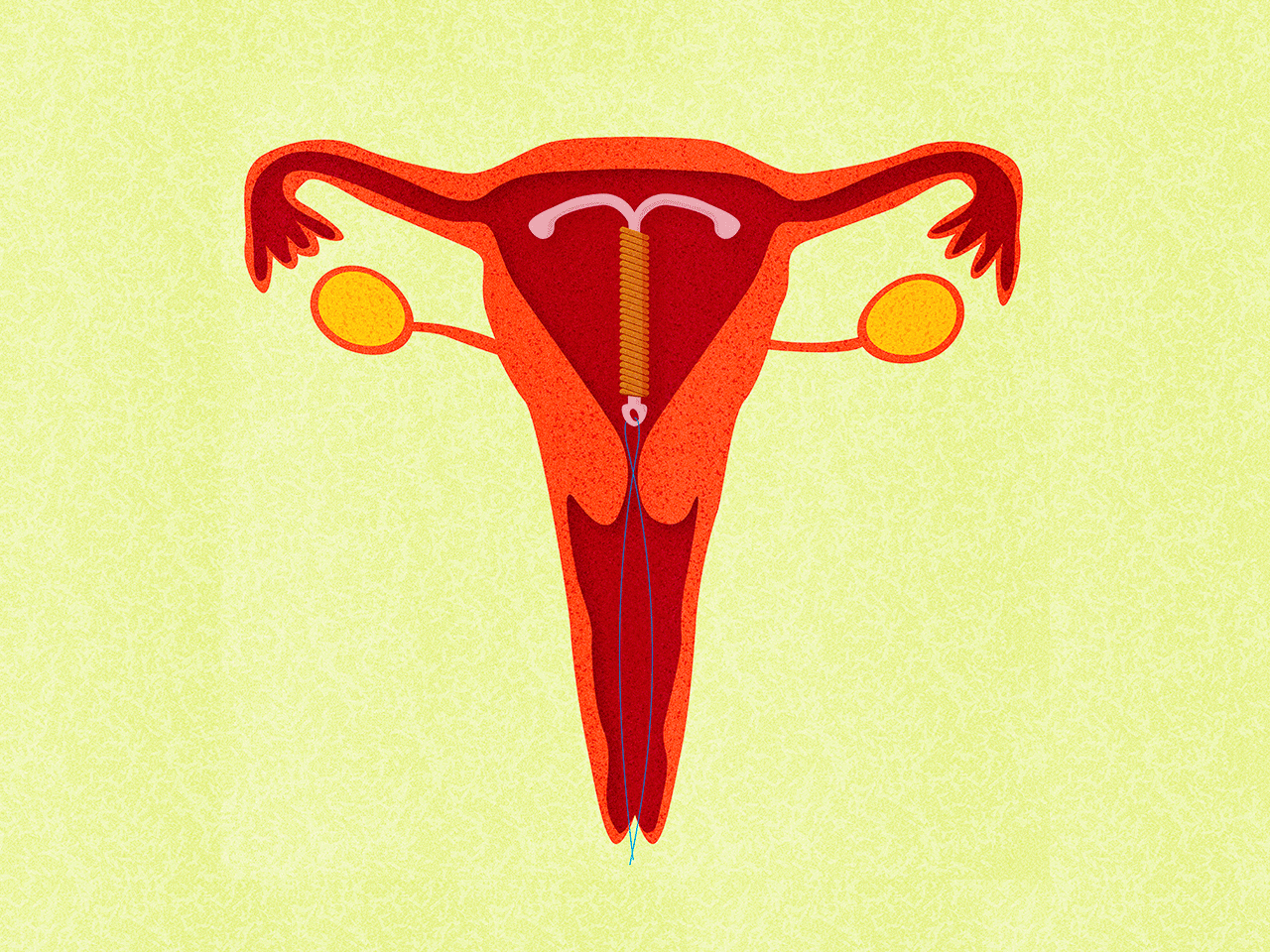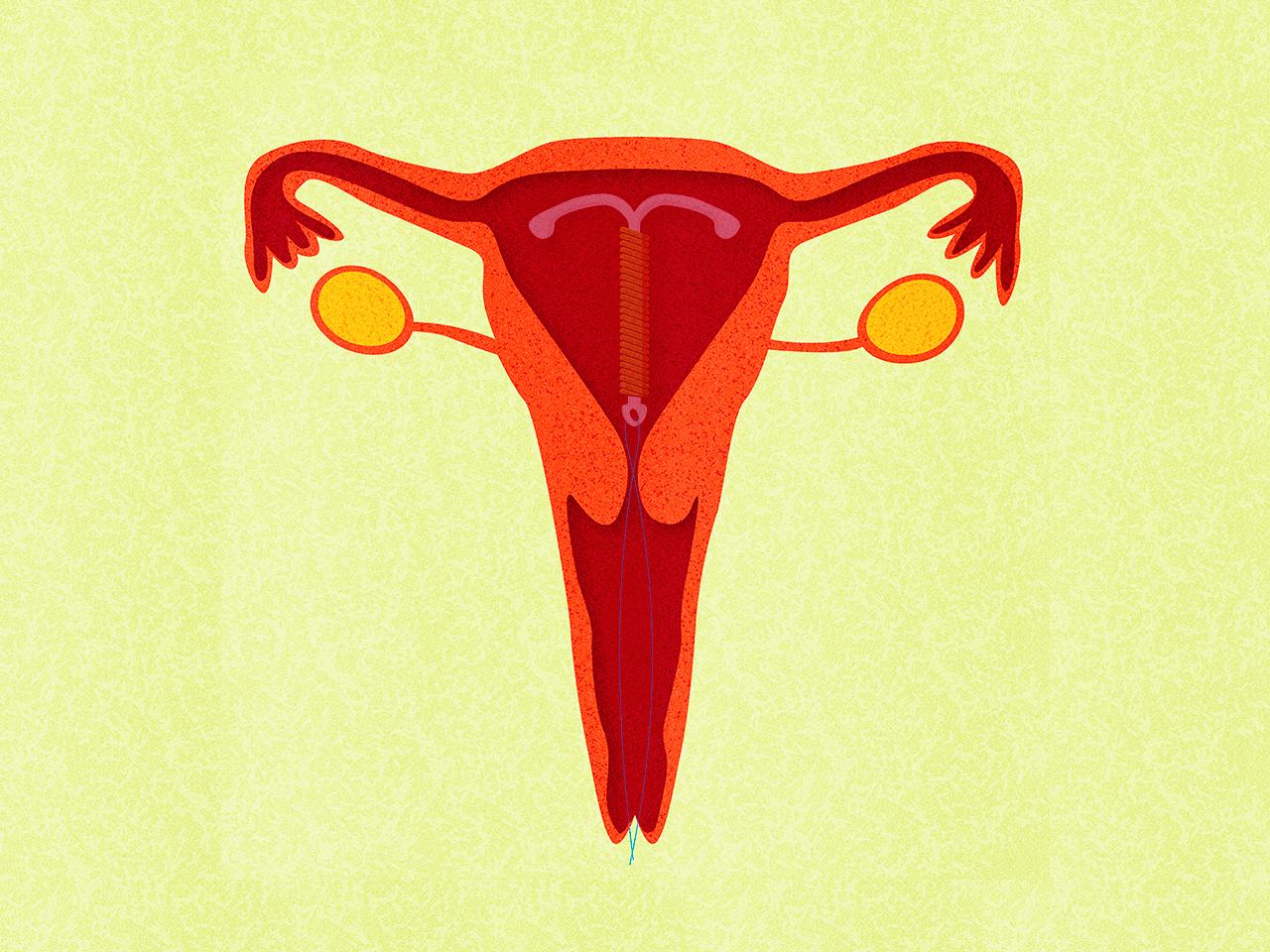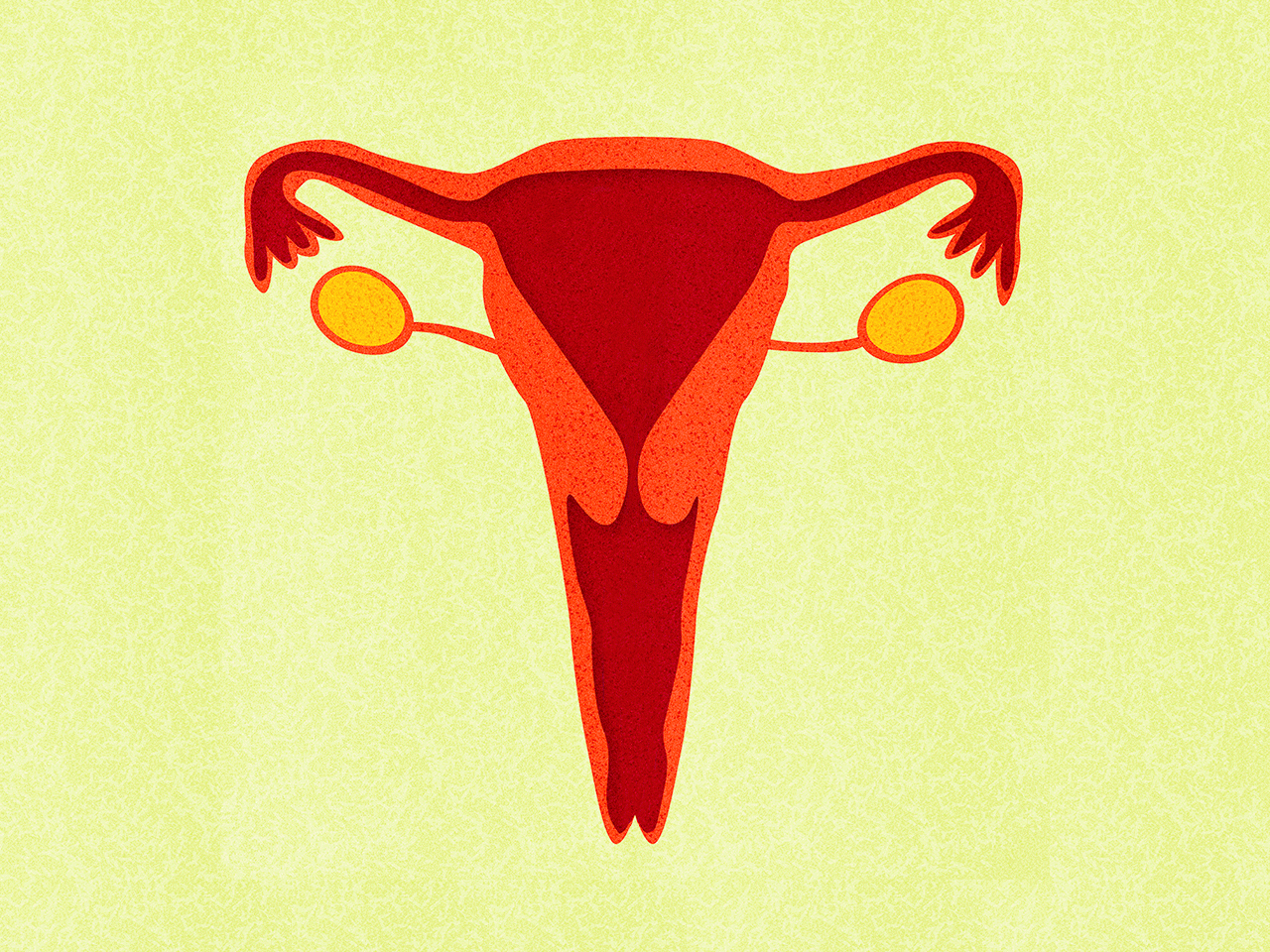
Внутриматочные гормональные системы содержат в своем составе гормон прогестин (левоноргестрел). Этот тип спирали также имеют Т-образный корпус. На корпусе расположена гормонально-эластомерная сердцевина, покрытая полупрозрачной мембраной, регулирующей поступление левоноргестрела в полость матки. Контрацептивный эффект таких спиралей обусловлен самой формой устройства и действием гормона прогестина.
Нефедов Глеб Александрович
акушер-гинеколог
Внутриматочная спираль обеспечивает надежную контрацепцию. Она устанавливается на срок до 5 лет. Мы используем разные спирали для рожавших и нерожавших женщин. Установку спирали проводит только врач гинеколог после предварительного обследования. Женщинам, которые выбрали этот метод контрацепции, стоит помнить, что защищает он лишь от нежелательной беременности, но не от половых инфекций.
Многие врачи являются сторонниками установки внутриматочной спирали, опираясь на следующие преимущества этого метода контрацепции:
- простота установки и удаления;
- длительный контрацептивный эффект;
- высокий процент эффективности;
- отсутствие необходимости ежедневного контроля;
- отсутствие негативного влияния гормональных препаратов, содержащих эстрогены;
- подходит для использования во время лактации;
- уже через 10 дней после снятия спирали полностью восстанавливается фертильность.

Для пациенток, страдающих ожирением, преимуществом метода является нейтральное воздействие ВМС на углеводный и жировой обмены, а также на систему гемокоагуляции и фибринолиза. Согласно рекомендациям ВОЗ медьсодержащие ВМС могут использоваться женщинами с ожирением без ограничений, вне зависимости от наличия макро- и микрососудистых осложнений, а также длительности заболевания Источник:
Макаров И.О. Контрацепция у пациенток с ожирением / И.О. Макаров, Е. И. Боровкова // Акушерство. Гинекология. Репродукция. — 2013. — Т.7. — № 1. — С. 36-40. .
К минусам можно отнести: невозможность защиты от венерических заболеваний, врачи не рекомендуют ставить спираль нерожавшим женщинам, остаётся возможность внематочной беременности, возможные межменструальные кровотечения, очень обильная потеря крови во время менструации.
Противопоказаниями к установке внутриматочной спирали могут стать:
- заболевания матки и ее шейки, особенно злокачественные опухоли;
- вагинальные инфекции;
- выделения из половых путей, обычно кровянистые, неустановленной этиологии;
- беременность;
- заболевания (воспалительные) органов малого таза;
- бесплодие.
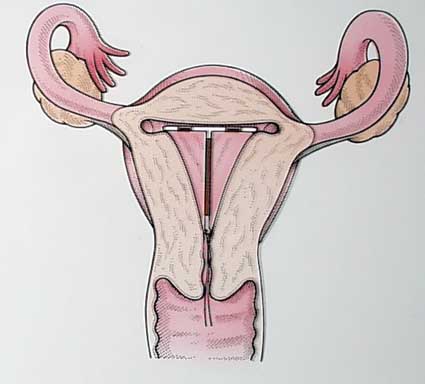
Не рекомендуется устанавливать внутриматочную спираль женщинам, имеющих рубцовую деформацию шейки матки после родов. Это связано с тем, что спираль не держится, поскольку сокращения матки выталкивают ее. Женщина может не почувствовать выпадения спирали, и может произойти зачатие Источник:
Махатова Г.М. Внутриматочная спираль как один из методов контрацепции /
Г.М. Махатова // Вестник хирургии Казахстана. — 2011. — № 3. — С. 74. .
Решившись на установку внутриматочной спирали, женщина должна пройти общеклиническое обследование, влагалищное исследование, УЗИ органов малого таза, расширенную кольпоскопию и бактериоскопическое исследование отделяемого из влагалища, осмотр молочных желез. Необходимо исключить онкологическую патологию органов женской половой сферы, беременность и инфекции, передающиеся половым путем. Все воспалительные гинекологические заболевания ко времени установки должны быть полностью излечены.
Установка внутриматочной спирали в «СМ-Клиника»
Если вы решились обезопасить себя, воспользовавшись внутриматочной спиралью, стоит обратиться к специалисту. Гинекологи «СМ-Клиника» успешно практикуют эту процедуру, используя только качественные спирали известных производителей.
Гинекологи «СМ-Клиника» успешно практикуют эту процедуру, используя только качественные спирали известных производителей.
Умелый персонал, новейшее оборудование, проверенные медикаменты и дневной стационар обеспечивают эффективное выполнение любой медицинской процедуры, в том числе постановки внутриматочной спирали.
Запишитесь на прием, и мы поможем вам!
Приём гинеколога в наших клиниках
Перед установкой внутриматочной спирали мне нужно сдавать какие-либо анализы? Сдавать анализы нужно обязательно. В список необходимых исследований входит:- бактериологическое исследование вагинальных и цервикальных мазков;
- обследование методом ПЦР на половые инфекции;
- клинический анализ крови и мочи;
- кровь на сифилис, ВИЧ, вирусные гепатиты В и С;
- расширенная кольпоскопия;
- УЗИ органов малого таза.
Какой тип внутриматочной спирали выбрать? По форме лучше использовать спирали с округлыми плечиками. В отличие от Т-образных, они не раздражают углы матки.
Используются в основном медьсодержащие или гормонопродуцирующие спирали. Их эффективность по данным ВОЗ составляет соответственно 93,8% и 99,8%. Таким образом, гормонопродуцирующие внутриматочные контрацептивы обеспечивают более надежную защиту от нежелательной беременности, поэтому они более предпочтительны.
Можно ли поставить спираль самостоятельно?
Ставить спираль самостоятельно нельзя. К тому же, это технически невозможно сделать в домашних условиях. Попытки самостоятельной установки не приведут к успеху, а лишь станут причиной травматизации половых органов и воспалительных процессов.
Источники:
- Макаров И.О. Контрацепция у пациенток с ожирением / И.О. Макаров, Е. И. Боровкова // Акушерство. Гинекология. Репродукция. — 2013. — Т.7. — № 1. — С. 36-40.
- Махатова Г.М. Внутриматочная спираль как один из методов контрацепции / Г.М. Махатова // Вестник хирургии Казахстана. — 2011. — № 3. — С. 74.
- Тихомиров А.Л. Внутриматочная гормональная контрацепция — локальная и логичная / А. Л. Тихомирова, С.И. Сарсания // Медицинский совет. — 2014. — С. 8-11.
Внутриматочная спираль (ВМС) — виды, установка, удаление в клинике ИнТайм
Надежность и длительный срок действия – одни из самых важных качеств внутриматочной спирали (ВМС). После установки ВМС применения дополнительных средств контрацепции не требуется. Процент эффективности варьируется от 97 до 99%. Срок эксплуатации ВМС, в зависимости от вида спирали, достигает 5-7 лет. Внутриматочные спирали не мешают естественной половой жизни, и не влияют на ее качество. А также не создают дискомфорта и в повседневной жизни.
Существует несколько видов спиралей – внутриматочная прогестерон-рилизинг система (ПРС), медная внутриматочная спираль, внутриматочная левоноргестрел-рилизинг система (ЛРС). Каждая из них имеет свои особенности.
Каждая из них имеет свои особенности.
Консультацию по выбору оптимального для вас типа ВМС вы можете получить в центре репродукции «ИнТайм». Опытные специалисты, гинекологи высшей и первой категории подберут для вас оптимальный вариант, пояснят принцип действия, безболезненно и быстро установят ВМС.
Отправьте заявкуна бесплатную консультацию
Мы осуществляем первую бесплатную консультацию для новых пациентов. Просто оставьте свои контактные данные, и мы свяжемся с вами в течение рабочего дня.
Если вы уже были нашим клиентом, вы можете воспользоваться этой формой для заказа обратного звонка.
Спасибо!
Ваша заявка принята.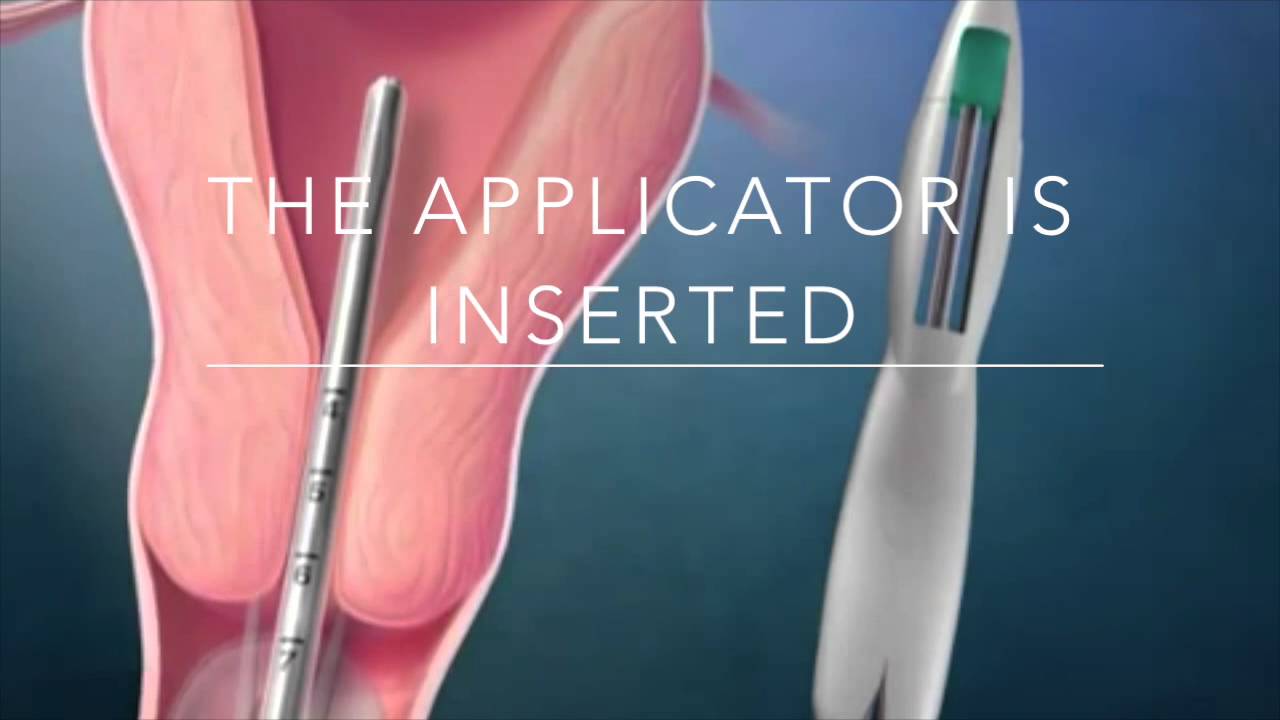
Мы свяжемся с вами в течение рабочего дня.
Почему стоит обратиться в нашу клинику
Компетентные сотрудники
Прием ведут опытные гинекологи, врачи высшей и первой категорий. Среди специалистов – кандидаты и доктора наук, профессора, академики. При подборе типа спирали компетентные специалисты учитывают физиологические особенности организма каждой пациентки и персональные пожелания.
Широкий выбор ВСМ разных типов
Внутриматочные спирали в нашей клинике представлены в нескольких модификациях, каждая из которых обладает своими особенностями и преимуществами. Для наших пациенток доступны лучшие внутриматочные спирали от ведущих мировых производителей (T de Oro 375, T de Plata 380 NOVAPLUS, Nova – T, «Мирена»).
Комфортные условия
Мы находимся в шаговой доступности от трех станций метро – Парк Культуры, Спортивная и Фрунзенская. К нам легко доехать и на личном транспорте – рядом Третье Транспортное кольцо. Вас ждут просторные кабинеты, доброжелательный персонал и отсутствие очередей – прием ведется по предварительной записи.
К нам легко доехать и на личном транспорте – рядом Третье Транспортное кольцо. Вас ждут просторные кабинеты, доброжелательный персонал и отсутствие очередей – прием ведется по предварительной записи.
Установка ВМС в клинике «ИнТайм»
Принцип действия
Пластиковые и металлические (золотые, серебряные) ВМС подавляют жизнедеятельность сперматозоидов, изменяя физиологическую трансформацию эндометрия и препятствуя вживлению оплодотворенной женской половой клетки. Из-за того, что ВМС стимулирует сокращения матки, эндометрий не успевает созреть к приему яйцеклетки, поэтому имплантация не происходит.
Гормоносодержащие спирали увеличивают вязкость цервикального секрета, что снижает миграцию мужских половых клеток через фаллопиевы трубы и матку и подавляет овуляцию.
Установка ВМС
Перед установкой спирали обязателен осмотр гинеколога, проведение расширенной кольпоскопии, УЗИ органов малого таза. Женщина сдает мазки на онкоцитологию и на флору из цервикального канала и влагалища.
Введение ВМС безболезненно и занимает несколько минут. Для минимальной травматизации цервикального канала спираль устанавливают на 1-2 день цикла, когда менструальные выделения наиболее обильны.
После установки ВМС врач назначает курс антибиотикотерапии. Через 4-5 дней женщина должна пройти первичный осмотр, и далее рекомендуется посещать гинеколога, как минимум, один раз в полгода.
Допускается введение ВМС поле неосложненного аборта. Также можно устанавливать спираль после родов, сразу после того как очистилась полость матки. Не рекомендуется установка ВМС нерожавшим женщинам.
Обращайтесь к опытным специалистам «ИнТайм»!
Наши специалисты индивидуально подберут для вас наиболее подходящую спираль, проведут установку или удаление ВМС по лояльным, доступным ценам в Москве.
Введение/или удаление внутриматочной спирали, кольца — цены на услуги гинекологии в Новокузнецке
Как действует ВМС и как часто ее нужно менять
Механические спирали просто препятствуют проникновению сперматозоидов в матку. Они не позволяют им оплодотворить яйцеклетку и эффективно защищают женщину от нежелательной беременности. Спираль назначают врачи-гинекологи изначально для местного лечения в случае различных гинекологических заболеваний, а уж защита от нежелательной беременности – это дополнительное преимущество такого лечения.
Они не позволяют им оплодотворить яйцеклетку и эффективно защищают женщину от нежелательной беременности. Спираль назначают врачи-гинекологи изначально для местного лечения в случае различных гинекологических заболеваний, а уж защита от нежелательной беременности – это дополнительное преимущество такого лечения.
Как правило, негормональную спираль устанавливают на три года. Это предотвращает возникновение такого заболевания, как эндометрит – воспаление полости матки. После установленного срока спираль подлежит удалению, после чего врач должен взять мазок-отпечаток на атипичные клетки. Через 7-10 дней будут результаты, которые необходимо будет узнать в клинике.
Повторно установить спираль можно по прошествии месяца, если все в порядке со здоровьем и женщина не планирует беременность. Это осуществляется после того, как пройдут месячные.
Противопоказания к введению спирали
Внутриматочные спирали не ставятся ни при каких обстоятельствах в таких случаях, как:
- наличие острых и хронических заболеваний органов малого таза;
- наличие злокачественных образований в шейке матки;
- наличие или планирование беременности;
- кровотечение при неясной этиологии;
- гиперполименорея;
- дисменорея;
- менометроррагия, если причина заболевания не установлена;
- миома матки;
- экстрагенитальные заболевания;
- частые экспульсии;
-
аллергия на материалы изготовления.
Подготовка и как происходит введение спирали
Когда у пациентки возникает желание использовать в качестве контрацептива внутриматочную спираль, необходимо записаться на осмотр к врачу-гинекологу. Специалист должен осуществить осмотр, чтобы определиться, какой тип спирали будет оптимальным в каждом конкретном случае. В некоторых случаях понадобится сделать анализы, например, мазок на цитологию шейки матки, обследование на ИППП, бактериологическое исследование мазка, ультразвуковое исследование органов малого таза.
Полная серия анализов не является обязательной, но при определенной клинической картине важно пройти полное обследование. Средство контрацепции должно максимально подходить женщине, чтобы его использование не имело негативных последствий.
Введение происходит на гинекологическом кресле, анестезия для этой манипуляции не требуется. Процедура полностью безболезненная, не вызывает неприятных ощущений, поэтому даже местной анестезии не понадобится.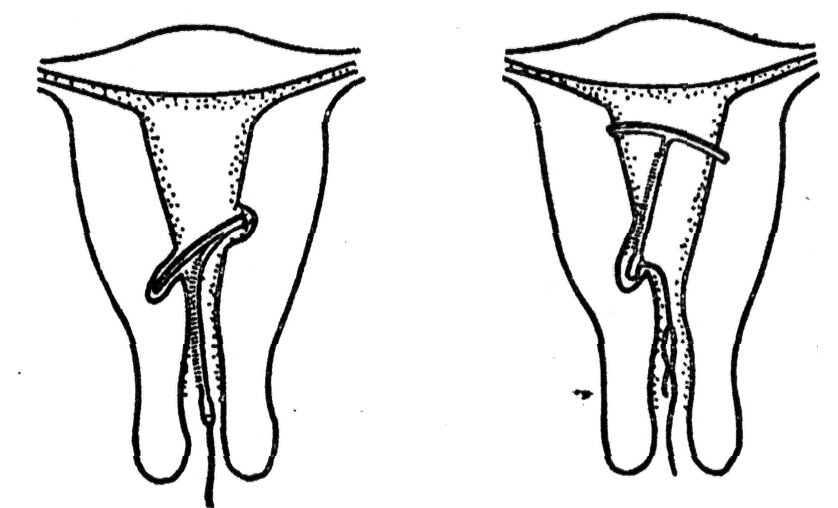 Через месяц необходимо прийти на контрольный осмотр, чтобы врач убедился в правильном положении контрацептива.
Через месяц необходимо прийти на контрольный осмотр, чтобы врач убедился в правильном положении контрацептива.
Причины удаления внутриматочной спирали
Обычной причиной удаления спирали является истечение срока, на который рекомендовано ее устанавливать. Однако извлечение может понадобиться и преждевременно. Это необходимо сделать в следующих случаях:
- если планируется беременность;
- есть желание женщины;
- появились боли;
- появились кровотечения, несущие угрозу жизни;
- начались воспалительные заболевания органов малого таза;
- появились злокачественные образования.
Подготовка и удаление спирали
Когда приходит пора удалять спираль, специалист производит осмотр и может назначить следующие виды исследования:
- УЗИ органов малого таза;
-
бактериологическое исследование мазка.

Если первоначально был установлен факт врастания спирали, дополнительно рекомендуются следующие исследования:
- анализ крови на гепатиты, ВИЧ, сифилис;
- общий и биохимический анализ крови.
Первые дни после завершения менструации являются наиболее благоприятными для удаления спирали. Важно заранее договориться с врачом-гинекологом на определенную дату. Удаление является безболезненным, как и установка. Анестезия не требуется. В случае, если ВМС вросла, используется гистероскоп. В таких условиях необходима местная анестезия или медикаментозный сон. Выскабливание эндометрия завершает такую манипуляцию.
Преимущества ВМС в клинике
Обращайтесь в клинику «Мать и дитя», если хотите воспользоваться всеми преимуществами такой контрацепции. Опытные специалисты изучат клиническую картину, чтобы определиться с лучшими вариантами установки спирали или кольца.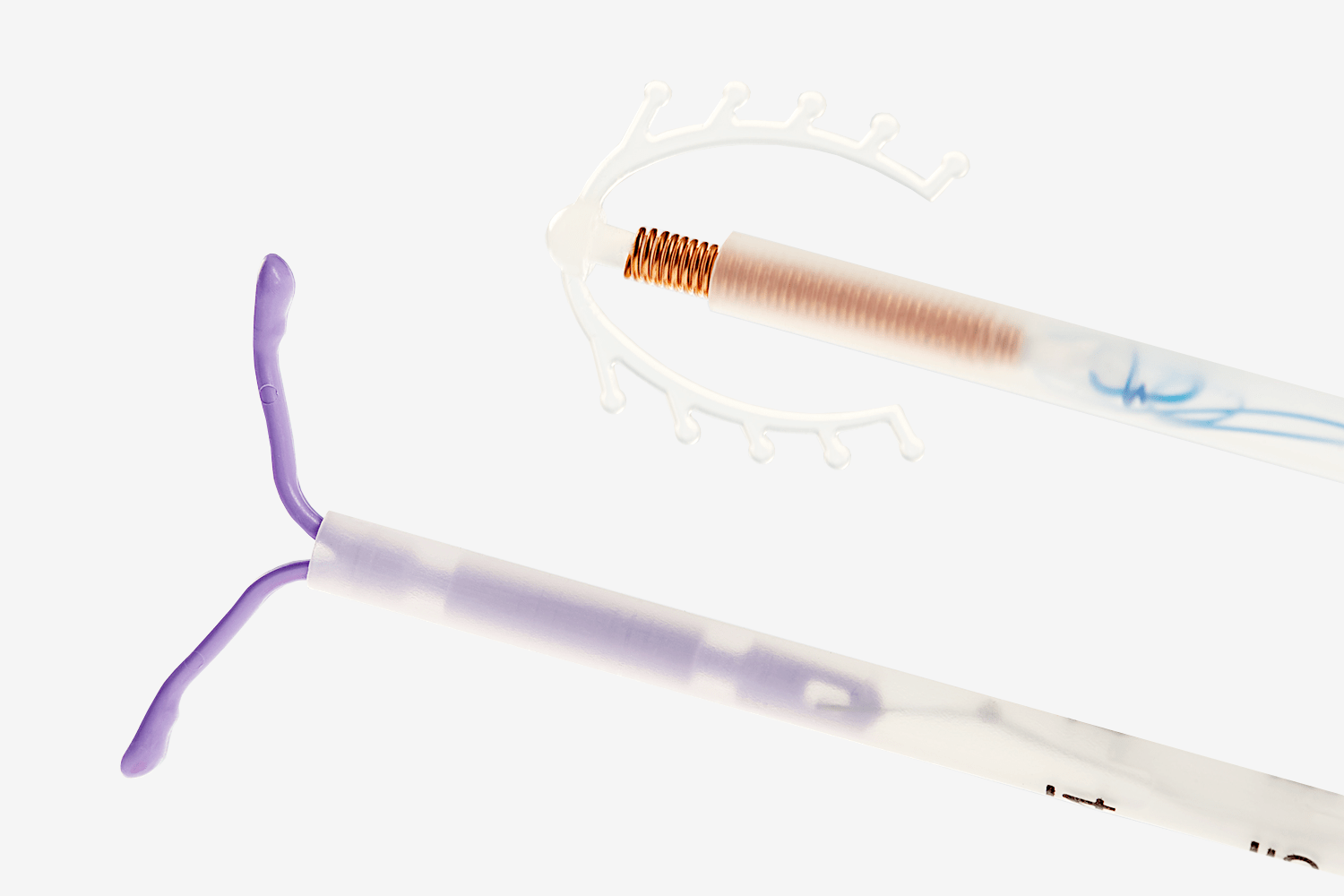 Безопасно и надежно предохраняться важно и нужно. Консультировать должны врачи с большим опытом, которые способны подсказать наиболее безопасные варианты.
Безопасно и надежно предохраняться важно и нужно. Консультировать должны врачи с большим опытом, которые способны подсказать наиболее безопасные варианты.
В нашей клинике вы сможете не только предохраняться от нежелательной беременности, но и запланировать долгожданную, наблюдать ее, чтобы весь процесс был безопасным. Мы предоставляем комплексные услуги, готовы обеспечить полноценный уход и максимум внимания каждой пациентке. С нашими врачами вы будете всегда уверены, что женское здоровье под контролем и для этого делается все необходимое!
Гинекология — Введение, удаление внутриматочной спирали
Установка внутриматочной спирали
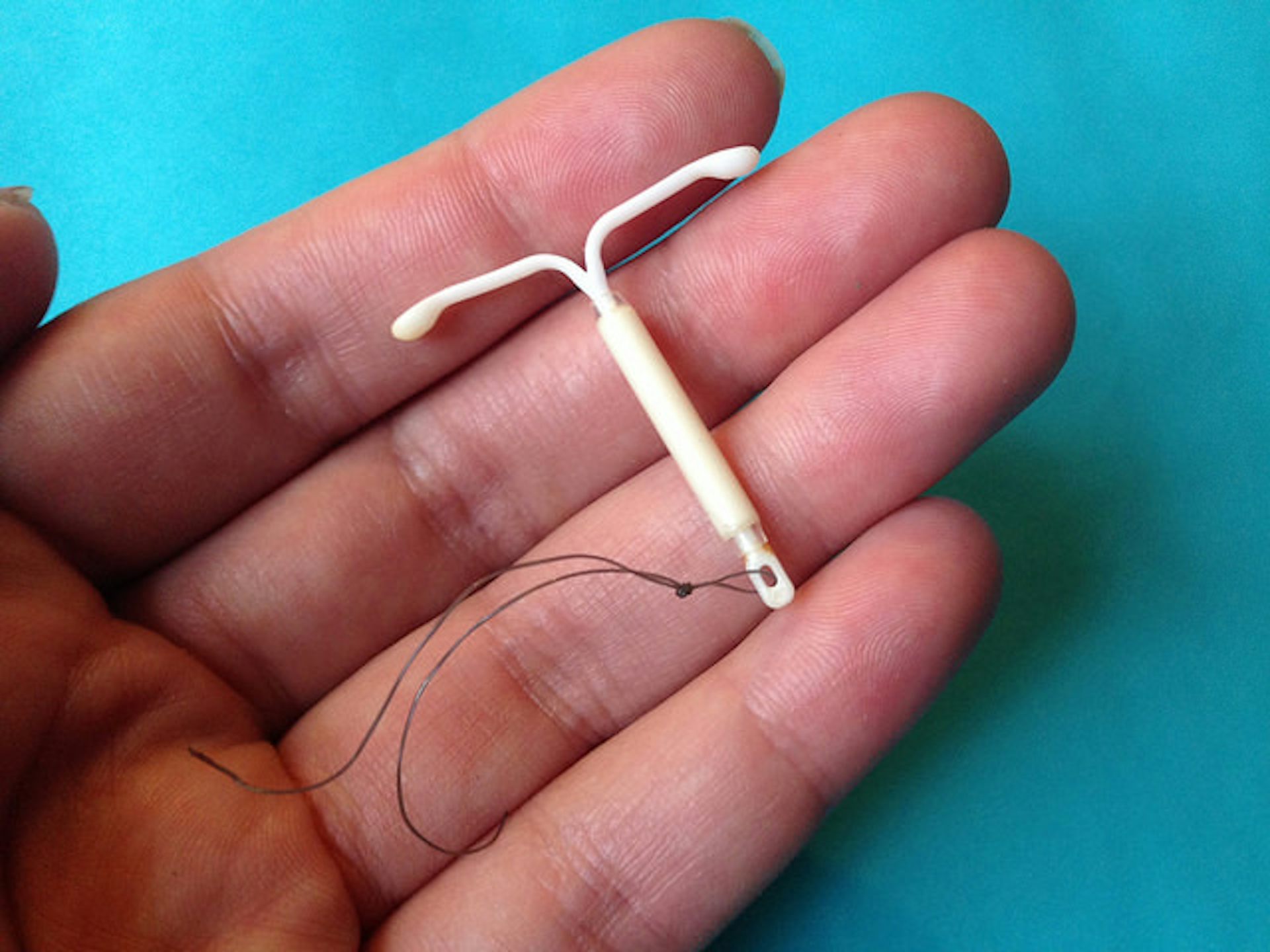
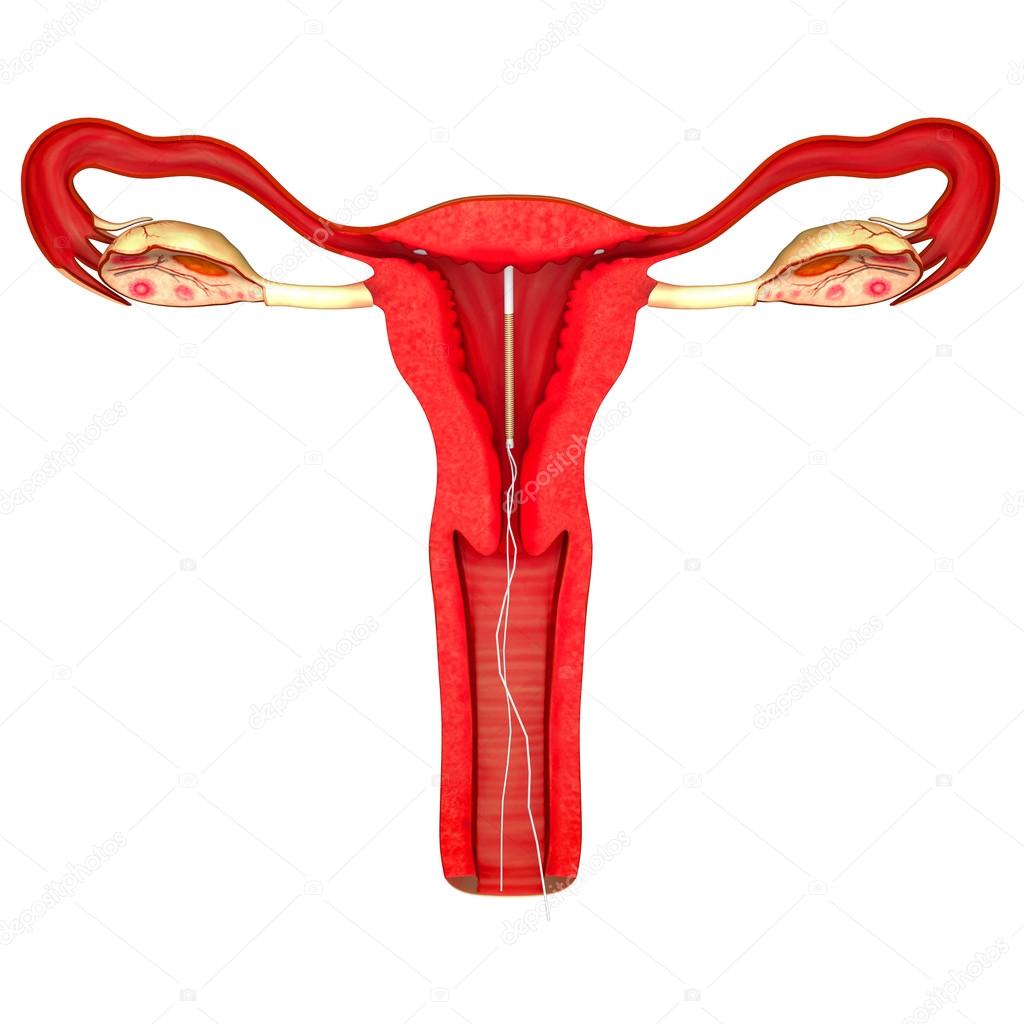
Внутриматочная спираль – это внутриматочный контрацептив, который устанавливается в полость матки женщины. Выглядит он как небольшая Т-образная пластиковая
палочка, на
основании которой спиралью намотаны медные волокна. Контрацепция осуществляется по такой схеме: установка внутриматочной спирали происходит как раз в том месте, в котором
оплодотворенная
яйцеклетка прикрепляется к матке, чтобы начать развитие эмбриона. Одновременно она является препятствием для продвижения сперматозоидов, и сокращает срок жизни яйцеклетки. То
есть,
сперматозоидов в полость матки проникает гораздо меньше. Двигательную функцию сперматозоидов угнетает медь, и она же препятствует появлению заражений и воспалений. А если
некоторые
все-таки смогли туда пробраться и даже смогли оплодотворить яйцеклетку, то ей просто негде начать разрастаться.
Одновременно она является препятствием для продвижения сперматозоидов, и сокращает срок жизни яйцеклетки. То
есть,
сперматозоидов в полость матки проникает гораздо меньше. Двигательную функцию сперматозоидов угнетает медь, и она же препятствует появлению заражений и воспалений. А если
некоторые
все-таки смогли туда пробраться и даже смогли оплодотворить яйцеклетку, то ей просто негде начать разрастаться.
Поставить спираль гинеколог может на срок от 3х до 5ти лет, а удаление внутриматочной спирали можно произвести в любой момент. Идеальное время для ее установки – 3-4 день после начала цикла. В это время шейка матки приоткрыта в связи с менструацией, и по этой же причине в эти дни почти полностью исключена вероятность забеременеть. При этом после ее установки неделю рекомендуется не вступать в половые контакты, а лучше подождать и 10 дней. Поставить спираль после родов можно только по истечении трех месяцев.
Перед тем, как поставить спираль, необходимо вылечиться от всех возможных патологий, которые могут присутствовать.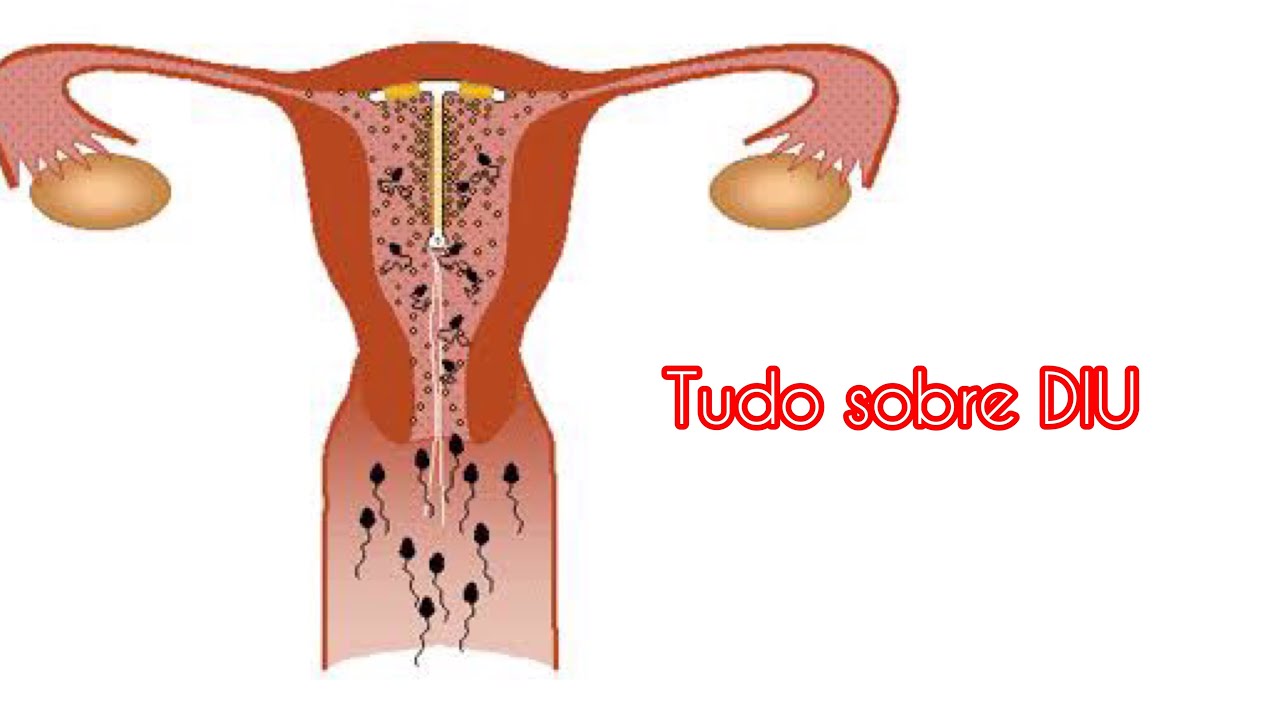 Поставить внутриматочную спираль можно, только если женщина
полностью
здорова в аспекте гинекологии. В ОК Центре в Харькове Вы также можете вылечить эрозию и дисбактериоз, которые препятствуют установке спирали. И раз в полгода
нужно появляться на осмотре
у гинеколога.
Поставить внутриматочную спираль можно, только если женщина
полностью
здорова в аспекте гинекологии. В ОК Центре в Харькове Вы также можете вылечить эрозию и дисбактериоз, которые препятствуют установке спирали. И раз в полгода
нужно появляться на осмотре
у гинеколога.
Преимущества и недостатки
Именно поэтому, что она находиться в полости матки, введение спирали чревато внематочной беременностью. Не пугайтесь, вероятность этого совсем не высока. Такой метод
контрацепции
препятствует беременности в 80% случаев. Только в оставшихся 20% процентах есть вероятность забеременеть, и эта беременность не обязательно должна быть внематочной. Самый
большой риск
забеременеть во время овуляции, то есть в тот самый 14 день. Поэтому можно завести календарь и высчитать его, для верности за день до и день после. В эти три дня лучше
применить
дополнительную контрацепцию или избегать половых актов. Такие меры предосторожности сведут риск беременности практически к нулю. А гормоносодержащие спирали и вовсе
предотвращают
овуляцию. Вместо медной проволоки они содержат контейнер, из которого постепенно выделяются гормоны. Они увеличивают процент защиты, но могут оказывать негативное воздействие
на
организм. Иногда женщины отмечают, что менструации длятся дольше и становятся более обильными. Но это обычно происходит, когда спираль установлена не верно. Могут возникнуть и
другие
осложнения, но они встречаются крайне редко, меньше чем в 1% проценте случаев.
А гормоносодержащие спирали и вовсе
предотвращают
овуляцию. Вместо медной проволоки они содержат контейнер, из которого постепенно выделяются гормоны. Они увеличивают процент защиты, но могут оказывать негативное воздействие
на
организм. Иногда женщины отмечают, что менструации длятся дольше и становятся более обильными. Но это обычно происходит, когда спираль установлена не верно. Могут возникнуть и
другие
осложнения, но они встречаются крайне редко, меньше чем в 1% проценте случаев.
Среди плюсов – спираль абсолютно не ощущается женщиной. Если ощущение инородного тела все-таки присутствует, значит, гинеколог установил ее неправильно. При половом акте мужчина
также
никоим образом не может ее почувствовать, если контрацептив установлен правильно. Если его ощущает один из партнеров, лучше сделать замену внутриматочной спирали. Еще одно
преимущество -
она не оказывает никакого воздействия на сам организм, как например, гормональные препараты. Плюс также и то, что обычно не требуется долго ждать восстановления репродуктивной
функции
при планировании беременности. Яйцеклетка может быть оплодотворена уже после первой менструации после удаления спирали. И, наверное, самое главное преимущество – это доступная
цена.
Плюс также и то, что обычно не требуется долго ждать восстановления репродуктивной
функции
при планировании беременности. Яйцеклетка может быть оплодотворена уже после первой менструации после удаления спирали. И, наверное, самое главное преимущество – это доступная
цена.
Цена внутриматочной спирали в Харькове
Бесспорный плюс поставить спираль – цена. Украина даже предлагает их как гуманитарную помощь. Но, это происходит редко, исключительно в городских больницах, в которых никогда нельзя быть уверенным в ее качестве, квалификации врача, и правильности его действий. Этот метод контрацепции более чем доступен, поэтому нет смысла экономить. Лучше обратиться в ОК Центр, где работают опытные профессионалы, а вероятность врачебной ошибки практически исключена.
Если Ваши средства ограничены, или просто Вам не свойственна расточительность, идеальным решением будет поставить спираль. Цена ее, для примера, равняется цену
гормональных препаратов на 2 месяца.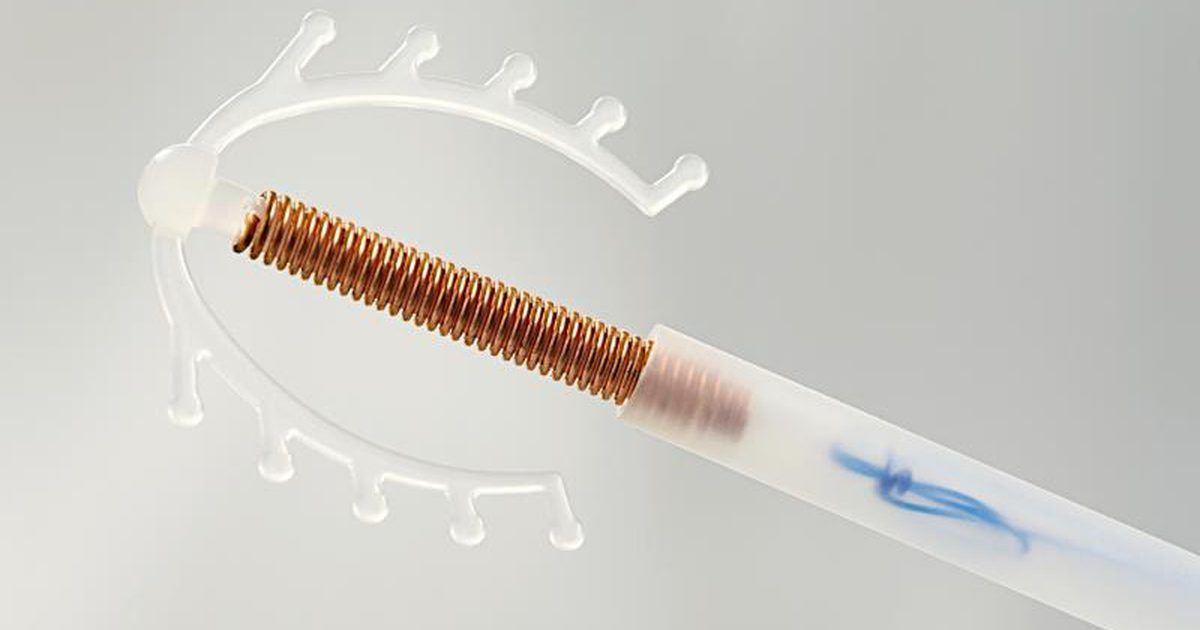 А прослужить она может несколько лет. Конечно, если Вы хотите поставить спираль после родов, цена будет
аналогична. Большинство женщин именно так и делают. Внутриматочные спирали Харьков предлагает чуть ли не на каждом шагу. Но ведь нужно позаботиться о своем здоровье, быть
уверенным и в
клинике, и в самом гинекологе. ОК Центр в Харькове имеет в своем штате только самых опытных врачей высокой квалификации. Вы можете не сомневаться в нашей
компетенции. Тем более что наши
цены очень доступны. Они указаны на сайте. Также на сайте вы найдете наши
контакты – адрес и телефон.
А прослужить она может несколько лет. Конечно, если Вы хотите поставить спираль после родов, цена будет
аналогична. Большинство женщин именно так и делают. Внутриматочные спирали Харьков предлагает чуть ли не на каждом шагу. Но ведь нужно позаботиться о своем здоровье, быть
уверенным и в
клинике, и в самом гинекологе. ОК Центр в Харькове имеет в своем штате только самых опытных врачей высокой квалификации. Вы можете не сомневаться в нашей
компетенции. Тем более что наши
цены очень доступны. Они указаны на сайте. Также на сайте вы найдете наши
контакты – адрес и телефон.
Внутриматочная спираль (ВМС) — установка в клинике Чудо-Доктор в Москве
Что такое внутриматочные контрацептивы и как они действуют?
К внутриматочным контрацептивам относят приспособления, которые вводят в полость матки с целью предотвращения беременности — это спирали и гормональные капсулы. Наиболее распространена внутриматочная спираль (ВМС).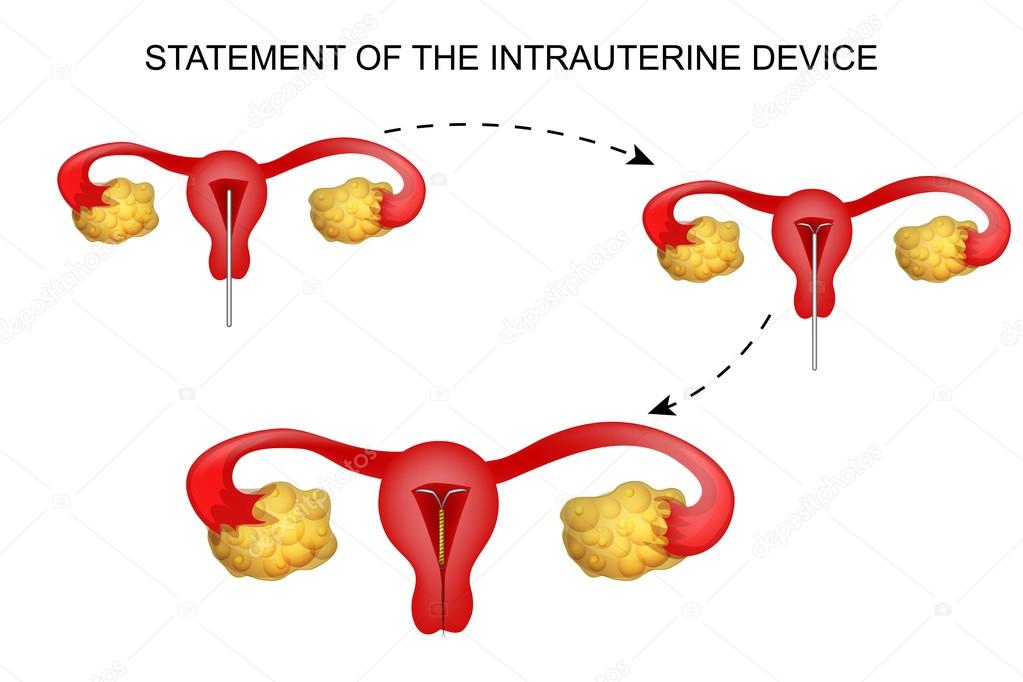 Внутриматочные спирали могут быть медикаментозные и немедикаметозные. Медикаментозные модели содержат медь и серебро и обладают комбинированным противозачаточным действием (сейчас используют только такие ВМС). Спираль представляет собой медную проволочку, обернутую вокруг эластичного стержня длиной 3-4 см, к которому прикреплены 2 нейлоновые нити. Спираль вводится врачом в полость матки, Оказавшись там, она препятствует наступлению беременности за счет присутствия инородного тела в матке и спермицидного эффекта меди. Спираль также усиливает сокращение матки и труб, вызывая специальное «асептическое» воспаление в полости матки, что создает неблагоприятные условия для наступления беременности.
Внутриматочные спирали могут быть медикаментозные и немедикаметозные. Медикаментозные модели содержат медь и серебро и обладают комбинированным противозачаточным действием (сейчас используют только такие ВМС). Спираль представляет собой медную проволочку, обернутую вокруг эластичного стержня длиной 3-4 см, к которому прикреплены 2 нейлоновые нити. Спираль вводится врачом в полость матки, Оказавшись там, она препятствует наступлению беременности за счет присутствия инородного тела в матке и спермицидного эффекта меди. Спираль также усиливает сокращение матки и труб, вызывая специальное «асептическое» воспаление в полости матки, что создает неблагоприятные условия для наступления беременности.
Гормональные внутриматочные контрацептивы — это различные приспособления, содержащие капсулы с гормональными препаратами, которые медленно высвобождаются в полости матки в течение 5 лет. Они оказывают местное действие на внутренние ткани матки, маточные трубы, слизистую оболочку шейки матки.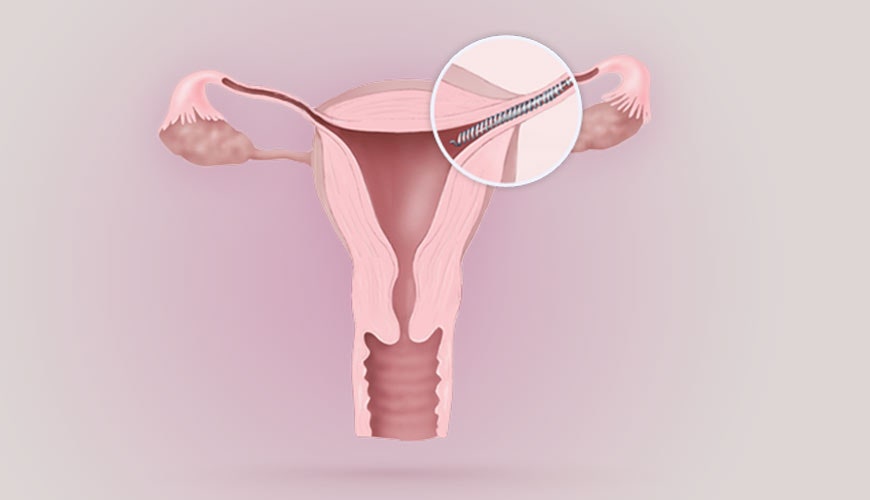
Кому могут быть рекомендованы внутриматочные спирали?
Внутриматочные спирали могут оказаться подходящим видом контрацепции для тех из нас, кто по какой-либо причине не могут принимать противозачаточные таблетки. Спирали рекомендуются женщинам, состоящим в браке и ведущим регулярную половую жизнь. При использовании спирали надо вести половую жизнь с постоянным половым партнером, так как спираль делает организм особо уязвимым для заболеваний, передающихся половым путем. Спираль может успешно применяться только в том случае, если вы практически здоровы, особенно с точки зрения гинекологии.
Когда нельзя применять внутриматочную спираль?
Для того, чтобы успешно применять спираль, вы должны быть гинекологически здоровы. Спираль можно вводить, если в течение 10-12 месяцев до этого вы не болели заболеваниями передающимися половым путем.
Противопоказания к применению спирали
- воспалительные заболевания органов малого таза после последней беременности; перенесенные ранее внематочные беременности;
- нарушение свертываемости крови;
- пороки сердца;
- хронические нарушения менструального цикла, в том числе очень болезненные или обильные месячные;
- высокий риск возникновения заболеваний, передающихся половым путем;
- любые заболевания, вредно действующие на иммунную систему или подавляющие сопротивляемость к инфекциям, например диабет или применение кортикостероидов.
- острые и хронические воспалительные заболевания наружных и внутренних половых органов;
- подтвержденная или предполагаемая беременность;
- маточные кровотечения неизвестного происхождения;
- подтвержденное или подозреваемое онкологическое заболевание гениталий; патологическое заболевание гениталий;
- патологические изменения матки, которые препятствуют успешному введению и эффективному использованию спирали.

Обследование перед введением спирали?
Прежде всего необходимо убедиться в том, что вы не больны инфекционно-воспалительным заболеванием. Поэтому врач обязан тщательно расспросить пациентку о состоянии ее здоровья, направить ее на анализы крови и мочи, взять специальные пробы ткани и бактериологические мазки. Для определения подходящего размера и модели спирали необходимо сделать ультразвуковое исследование матки.
Как и когда вводят спираль?
Эту процедуру осуществляют только в медицинских учреждениях.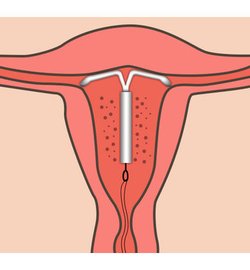 Обычно спираль упакована в стерильные пакеты со специальными приспособлениями, что позволяет ввести спираль в полость матки на гинекологическом кресле за 2-3 минуты. Шейки матки предварительно смазывают дезинфицирующим и обезболивающим растровом. При введении спирали огромное значение имеет соблюдение стерильности. Большинство женщин переносят эту манипуляцию легко и безболезненно. Некоторые испытывают легкую боль, дурноту и головокружение. Реже введение спирали сопровождается кратковременной сильной болью, которая может вызвать обморочное состояние, тошноту и т.д. Эти ощущения быстро проходят. Лучший срок для постановки спирали — последние дни менструаций, первые 6 недель после родов, спираль также может быть введена во время медицинского аборта.
Обычно спираль упакована в стерильные пакеты со специальными приспособлениями, что позволяет ввести спираль в полость матки на гинекологическом кресле за 2-3 минуты. Шейки матки предварительно смазывают дезинфицирующим и обезболивающим растровом. При введении спирали огромное значение имеет соблюдение стерильности. Большинство женщин переносят эту манипуляцию легко и безболезненно. Некоторые испытывают легкую боль, дурноту и головокружение. Реже введение спирали сопровождается кратковременной сильной болью, которая может вызвать обморочное состояние, тошноту и т.д. Эти ощущения быстро проходят. Лучший срок для постановки спирали — последние дни менструаций, первые 6 недель после родов, спираль также может быть введена во время медицинского аборта.
Осложнения при применении спирали
Частым осложнением является выпадение спирали из полости матки. Чаще всего это происходит в первые же часы после введения спирали или в первые три месяца ее использования. Если спираль остается в матке после этого периода, то специалисты говорят, сто она «прижилась».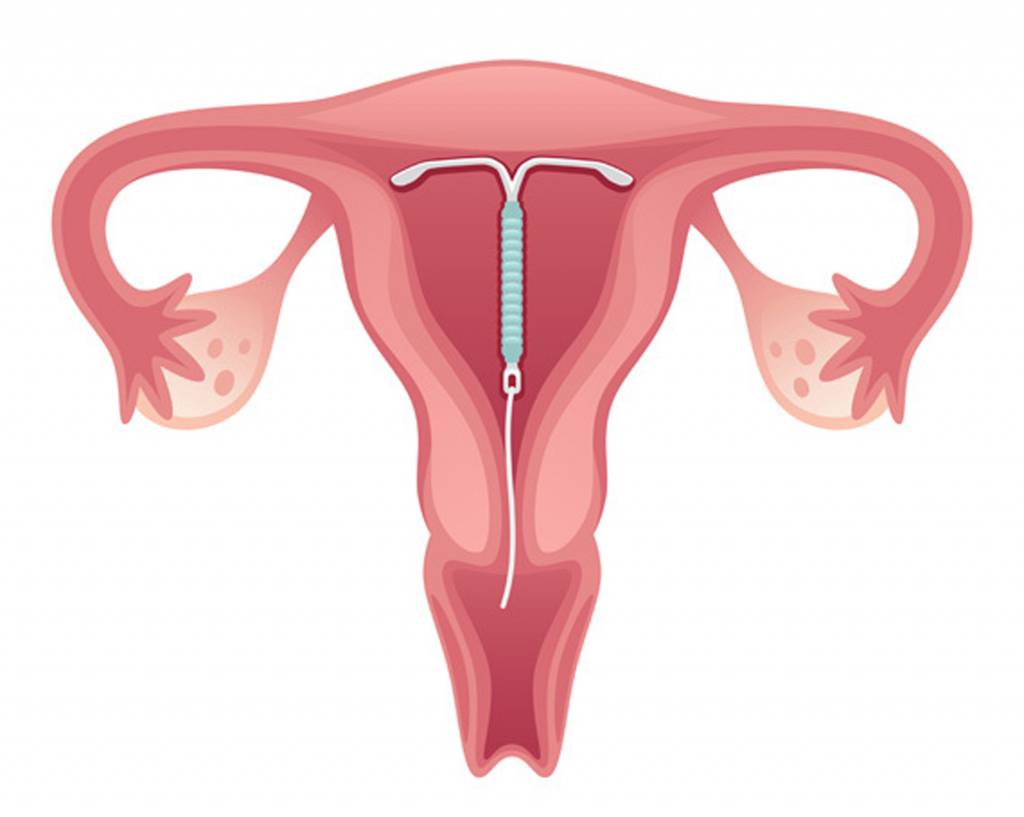 Само по себе выпадение спирали не является опасным, важно вовремя заметить это и обратиться к врачу, который подберет более подходящий размер и модель спирали.
Само по себе выпадение спирали не является опасным, важно вовремя заметить это и обратиться к врачу, который подберет более подходящий размер и модель спирали.
В течение первых 3-8 месяцев ваш организм «привыкает» к спирали. Именно в этот период могут возникнуть различные осложнения: обостряются воспалительные заболевания половых органов, проявляются скрытые инфекции мочеполовых путей. Поэтому при появлении таких симптомов, как зуд и жжение во влагалище, боли в пояснице и внизу живота, повышение температуры, общее недомогание, озноб, необходимо обратиться к врачу и начать лечение болезни.
Частыми осложнениями при введении спирали бывают различные маточные кровотечения: от умеренных кровянистых выделений между менструациями до обильных изнурительных кровотечений во время месячных. В таких случаях надо провести индивидуальное лечение. Если же кровотечение принимает постоянный характер и поддается лечению лекарственными препаратами, то возникает необходимость удалить спираль.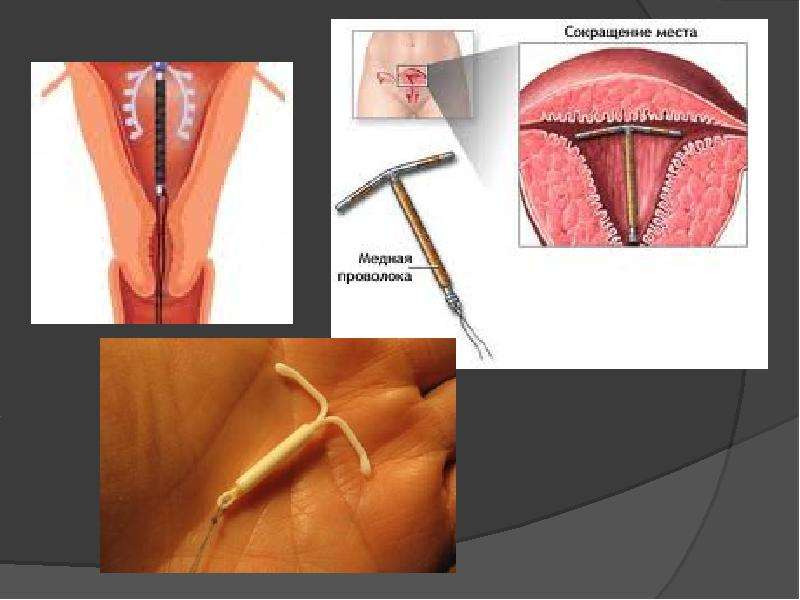 Если введение спирали чрезвычайно болезненно или очень затруднительно, то это весомый повод для того, чтобы отказаться от внутриматочного контрацептива.
Если введение спирали чрезвычайно болезненно или очень затруднительно, то это весомый повод для того, чтобы отказаться от внутриматочного контрацептива.
Преимущества
Эффективность спирали — 98%. Она рассчитана на длительные сроки применения: от 2 до 5 лет. После введения обычно нет необходимости в специальном медицинском наблюдении. Спираль не оказывает влияния на весь организм. Способность к зачатию восстанавливается в течении 1-2 менструальных циклов. Показаны женщинам старше 35 лет и для тех, кому противопоказаны противозачаточные таблетки. Спирали доступны по цене, процедура их постановки не занимает много времени.
Может ли спираль повредить органы партнера во время полового акта?
Даже если спираль выпадет из матки во время коитуса (чего практически не отмечается), то она не может травмировать половой член, так как имеет закругленную и обтекаемую форму.
Спиральные артерии матки при беременности человека: факты и противоречия
Резюме
Спиральные артерии матки играют жизненно важную роль в снабжении питательными веществами плаценты и плода, и для этой цели они трансформируются в сильно расширенные сосуды под действием вторгающегося трофобласта.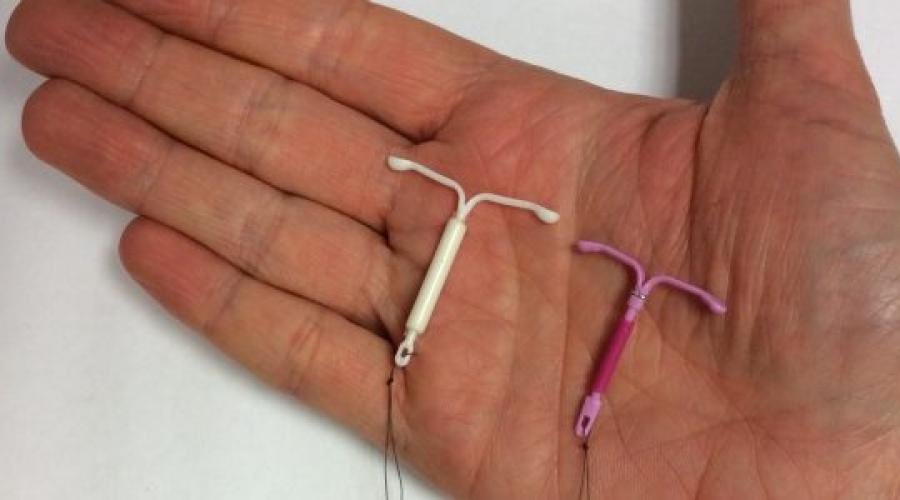 (физиологическое изменение). Знание механизмов этих изменений важно для лучшего понимания преэклампсии и других осложнений беременности, которые свидетельствуют о неполном ремоделировании спиральной артерии.Споры по поводу различных этапов этих физиологических изменений все еще существуют, и некоторые из этих разногласий освещены в этом обзоре, предлагая направления для дальнейших исследований. Во-первых, лучшее определение степени ремоделирования децидуальной оболочки по сравнению с трофобластом может помочь разработать более адекватную терминологию. Другими спорными проблемами являются закупорка сосудов и их связь с кислородом, инвазия трофобластов снаружи или изнутри сосудов (интравазация по сравнению с экстравазацией), влияние гемодинамики на эндоваскулярную миграцию, замещение артериальных компонентов трофобластом, механизмы восстановления материнской ткани. и роль естественных киллеров (NK) матки.Некоторые из этих особенностей могут быть нарушены при осложненной беременности, включая раннее ремоделирование сосудов, связанное с децидуальной оболочкой, закупорку сосудов и гемодинамику.
(физиологическое изменение). Знание механизмов этих изменений важно для лучшего понимания преэклампсии и других осложнений беременности, которые свидетельствуют о неполном ремоделировании спиральной артерии.Споры по поводу различных этапов этих физиологических изменений все еще существуют, и некоторые из этих разногласий освещены в этом обзоре, предлагая направления для дальнейших исследований. Во-первых, лучшее определение степени ремоделирования децидуальной оболочки по сравнению с трофобластом может помочь разработать более адекватную терминологию. Другими спорными проблемами являются закупорка сосудов и их связь с кислородом, инвазия трофобластов снаружи или изнутри сосудов (интравазация по сравнению с экстравазацией), влияние гемодинамики на эндоваскулярную миграцию, замещение артериальных компонентов трофобластом, механизмы восстановления материнской ткани. и роль естественных киллеров (NK) матки.Некоторые из этих особенностей могут быть нарушены при осложненной беременности, включая раннее ремоделирование сосудов, связанное с децидуальной оболочкой, закупорку сосудов и гемодинамику.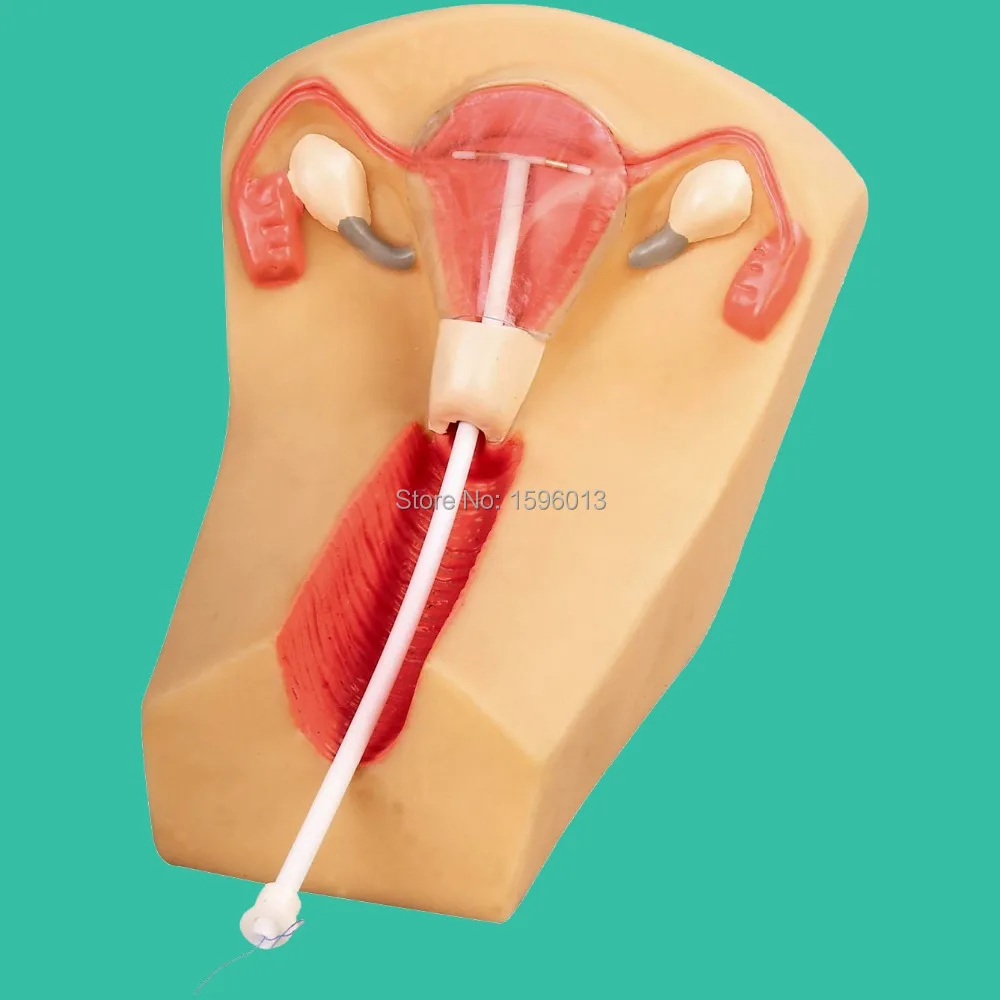 Гипервоспалительное состояние преэклампсии может быть причиной васкулопатий, таких как острый атероз, хотя общее влияние таких поражений на функцию плаценты далеко не ясно. Некоторые особенности человеческого плацентарного ложа отражаются процессами у других видов с гемохориальной плацентой, и изучение таких моделей может помочь пролить свет на плохо изученные аспекты человеческой плацентации.
Гипервоспалительное состояние преэклампсии может быть причиной васкулопатий, таких как острый атероз, хотя общее влияние таких поражений на функцию плаценты далеко не ясно. Некоторые особенности человеческого плацентарного ложа отражаются процессами у других видов с гемохориальной плацентой, и изучение таких моделей может помочь пролить свет на плохо изученные аспекты человеческой плацентации.
Ключевые слова
Спиральные артерии
Инвазия трофобластов
Ремоделирование сосудов
Человек
Преэклампсия
Децидуализация
NK-клетки матки
Рекомендуемые статьи 9Citing статьи Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
Гормональные спирали противостоят раку матки
Прошло почти десять лет с тех пор, как исследователи из Университета Тромсё (UiT) впервые обнаружили, что женщины, которые использовали гормональные контрацептивы, также получают преимущества профилактических эффект против рака матки.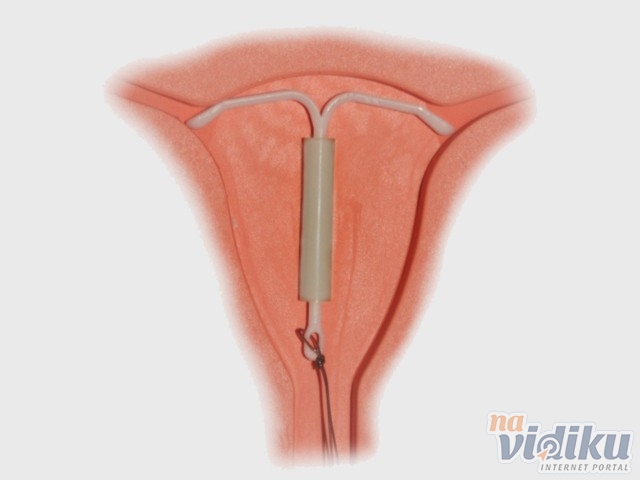
Энн Орбо, профессор гинекологии в Университете штата Калифорния, сообщает, что ранние стадии рака исчезают у всех женщин в группе, получавшей гормональную спираль Мирена в течение шести месяцев, а затем наблюдалась в течение двух лет.
У женщин не было никаких признаков клеточных изменений, которые могут развиться в злокачественные опухоли, пока они использовали спираль.
Спираль превосходит планшеты
Гормональная спираль превосходит лечение таблетками.Результаты благоприятны, а тенденции очевидны: все женщины в группе, использовавшие спираль, откликнулись.
«Это означает, что предварительные стадии рака матки полностью исчезли с помощью спирали», — говорит Эрбо, первый автор нового исследования, которое вскоре будет опубликовано в международном журнале.
Требуется национальное руководство
Всего 170 женщин, у всех из которых развились предварительные стадии рака матки, были обследованы в ходе случайного контрольного исследования.
Это метод исследования, который считается наиболее подходящим для изучения и сравнения эффектов и форм лечения.
Исследователи никогда раньше не оперировали с таким длительным периодом наблюдения в два года в исследованиях лечения гормональной спиралью по сравнению с лечением таблетками.
Гормональная спираль, Т-образный кусок пластика, помещенный внутрь матки. Кусочек пластика содержит медь или синтетический гормон прогестерон, предотвращающий беременность. (Фото: iStockphoto)
Новые результаты могут привести к новым национальным рекомендациям по лечению пациентов с ранними стадиями рака матки.
В настоящее время в Норвегии отсутствуют общие национальные распорядки. Прогестерон в таблетированной форме долгое время был консервативным методом лечения, но гистерэктомия также преобладала, даже среди молодых женщин.
«Я бы сказал, что слишком много женщин перенесли гистерэктомию, — объясняет профессор. — Наши результаты показывают, что в будущем нам нужно работать по-другому. Выбирая людей с самым высоким риском и снабжая их гормональными спиралями, мы можем сократить количество случаев рака и операций.”
Выбирая людей с самым высоким риском и снабжая их гормональными спиралями, мы можем сократить количество случаев рака и операций.”
Возврат при снятии спирали
Орбо испытал, что гормональная спираль заставляет опасные мутации отступать на клеточном уровне. Пока неясно, какой механизм срабатывает в этом контексте после того, как противозачаточное устройство введено в действие.
Современное лечение рака матки
Самым распространенным методом лечения было хирургическое удаление матки, яичников и маточных труб.
Иногда также удаляются лимфатические узлы в тазу.
В некоторых случаях при метастазах в яичники их тоже удаляют.
Если опухоль распространилась далеко в стенку матки или за ее пределы, пациентка после операции получает лучевую терапию.
Если рак матки находится на начальной стадии, обычно удаляется только матка.
Источник: Норвежское онкологическое общество, Gynkreftforeningen, Anne Ørbo.
Результаты показывают, что положительный эффект гормональной спирали длится до тех пор, пока женщина сохраняет гормональную спираль в своей матке.
Профессор считает, что начальные стадии рака либо обращены вспять, либо мутации скрываются и остаются пассивными, пока активна гормональная спираль. Гормональные спирали необходимо заменить после пяти лет использования.
«Мы указываем, что при удалении спирали возвращается около 50 процентов раковых заболеваний на начальной стадии. Похоже, что женщины, особенно те, кто находится в группах риска по этому типу рака, должны постоянно использовать эту спираль для достижения защитного эффекта », — говорит Орбо, возглавляющий исследовательскую группу по гинекологическому раку в Тромсё.
Отстает от рака шейки матки
Почти все случаи рака матки развиваются на начальных стадиях. Около 20 процентов этих заболеваний на ранней стадии перерастают в рак.
Рак матки был вытеснен раком шейки матки, который представляет собой совершенно другое заболевание. Вакцина против ВПЧ была в центре внимания как профилактическая мера от последнего типа рака.
Вакцина против ВПЧ была в центре внимания как профилактическая мера от последнего типа рака.
Кроме того, рак шейки матки может поражать женщин даже в подростковом возрасте, в то время как рак матки обычно не развивается, пока женщинам не исполнится 60 лет.
Тем не менее, все больше женщин в Норвегии заболевают раком матки, и это уносит больше жизней. С 1999 по 2009 год число случаев рака матки увеличилось с 492 до 696.
Для сравнения, ежегодно у 250-300 женщин диагностируется рак шейки матки.
Ожирение — самый большой фактор риска
Гормональная спираль работает, высвобождая небольшие дозы синтетического гормона, который имеет эффект прогестерона.
Хотя повышенный уровень женского гормона эстрогена увеличивает риск рака матки, прогестерон помогает внутренней слизистой оболочке (эндометрию) сокращаться и защищает ее от развития рака.
Поскольку рак матки начинается в самой матке и зависит от эстрогена, любые факторы, повышающие уровень эстрогена в организме, также могут повысить риск рака матки.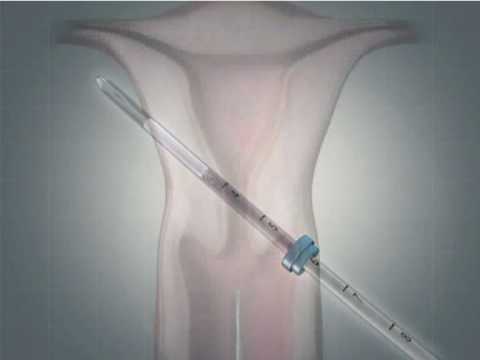
Самый большой фактор риска связан с избыточным весом. У тучной женщины риск заболеть раком матки в три раза выше.
«Нарушения обмена веществ, такие как диабет, также повышают риск, как и высокое кровяное давление, а также если женщины принимают эстроген в качестве гормональной добавки или антиэстрогены в результате рака груди», — говорит Орбо.
Несколько побочных эффектов
Профессор говорит, что только один процент женщин в новом исследовании сообщили о побочных эффектах, таких как нерегулярные, а иногда и чрезмерные менструальные кровотечения.
Всего в новом исследовании приняли участие 170 женщин из восьми больниц и девяти гинекологических центров страны. Он проводился независимо от всех фармацевтических компаний.
Одна группа использовала гормональную спираль, а две группы использовали таблетки с разными дозировками.
Исследователи засвидетельствовали обратное развитие ранних стадий рака среди всех пользователей спирали. В группе таблеток с более низкой дозировкой этого улучшения здоровья достигла только половина.
В группе таблеток с более низкой дозировкой этого улучшения здоровья достигла только половина.
В группе таблеток, принимавших более высокую дозу гормона, было много женщин, которые прекратили лечение из-за побочных эффектов. Поэтому Орбо и ее коллеги считают это наименее полезным методом лечения.
————————————————- —
Прочтите эту статью на норвежском языке на сайте forskning.нет
Перевод: Гленн Остлинг
Децидуальная ниша матки модулирует прогрессивную дедифференцировку гладкомышечных клеток сосудов спиральной артерии во время беременности человека † | Биология размножения
Аннотация
Ремоделирование спиральной артерии матки (SPA) является важным событием во время беременности для обеспечения достаточного кровоснабжения границы раздела матери и плода и удовлетворения потребностей растущего плода. Наряду с этим процессом, динамическое изменение и судьба гладкомышечных клеток сосудов спиральных артерий (SPA – VSMCs) долгое время оставались предметом споров.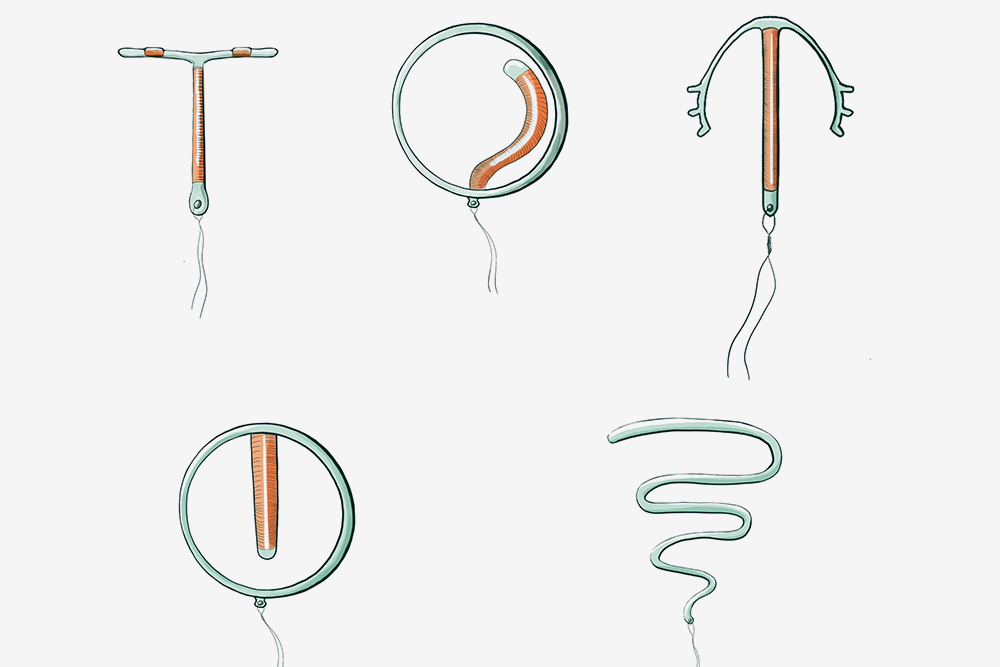 В настоящем исследовании мы проанализировали клеточные особенности SPA – VSMC на разных этапах ремоделирования сосудов у человека на ранних сроках беременности и продемонстрировали прогрессивное морфологическое изменение SPA – VSMC в немоделированных (Un-Rem), ремоделированных и полностью ремоделированные (Fully-Rem) стадии, указывающие на независимую от вневорсинчатого трофобласта (EVT) и EVT-зависимую фазы дедифференцировки SPA-VSMC. Эксперименты in vitro на клеточной линии VSMC выявили эффективную роль децидуальных стромальных клеток, децидуальных естественных клеток-киллеров (dNK), децидуальных макрофагов и EVT в индукции дедифференцировки VSMC.Важно отметить, что потенциальная трансформация VSMC в CD56 + dNK была показана с помощью иммунофлуоресценции ДНК in-situ гибридизации-лигирования близости и анализа иммунопреципитации хроматина для модификации h4K4dime в промоторной области тяжелой цепи миозина 11 ( MYh21 ). Полученные данные ясно иллюстрируют каскадную регуляцию прогрессивной дедифференцировки SPA-VSMCs с помощью множественных типов клеток в децидуальной нише матки и предоставляют новые доказательства для выявления предназначения SPA-VSMCs во время ремоделирования сосудов.
В настоящем исследовании мы проанализировали клеточные особенности SPA – VSMC на разных этапах ремоделирования сосудов у человека на ранних сроках беременности и продемонстрировали прогрессивное морфологическое изменение SPA – VSMC в немоделированных (Un-Rem), ремоделированных и полностью ремоделированные (Fully-Rem) стадии, указывающие на независимую от вневорсинчатого трофобласта (EVT) и EVT-зависимую фазы дедифференцировки SPA-VSMC. Эксперименты in vitro на клеточной линии VSMC выявили эффективную роль децидуальных стромальных клеток, децидуальных естественных клеток-киллеров (dNK), децидуальных макрофагов и EVT в индукции дедифференцировки VSMC.Важно отметить, что потенциальная трансформация VSMC в CD56 + dNK была показана с помощью иммунофлуоресценции ДНК in-situ гибридизации-лигирования близости и анализа иммунопреципитации хроматина для модификации h4K4dime в промоторной области тяжелой цепи миозина 11 ( MYh21 ). Полученные данные ясно иллюстрируют каскадную регуляцию прогрессивной дедифференцировки SPA-VSMCs с помощью множественных типов клеток в децидуальной нише матки и предоставляют новые доказательства для выявления предназначения SPA-VSMCs во время ремоделирования сосудов.
Введение
При беременности у человека ремоделирование спиральных артерий матки (SPA) клетками вневорсинчатого трофобласта (EVT) широко происходит на ранних и средних сроках беременности. Благодаря этому просвет артерии увеличивается, а сопротивление кровотоку в артерии прогрессивно снижается [1]. Этот процесс обеспечивает соответствующий приток материнской крови к границе раздела матери и плода для удовлетворения растущих потребностей растущего плода. Неполное ремоделирование SPA тесно связано с такими осложнениями беременности, как выкидыш, преэклампсия и задержка роста плода [2, 3].Поэтому всестороннее понимание механизмов ремоделирования SPA имеет важное значение для разработки эффективных стратегий вмешательства для этих клинических симптомов.
В то время как накапливающиеся исследования сосредотачиваются на роли EVT в ремоделировании SPA, критическая начальная стадия, включая физиологические изменения в эндотелии и гладких мышцах сосудов, происходит до инвазии EVT. Исследование продемонстрировало вакуолизацию эндотелиальных клеток и набухание некоторых отдельных SPA гладкомышечных клеток сосудов (SPA – VSMC) на очень ранней стадии процесса ремоделирования [4].Инвазия EVT связана с дальнейшей дезорганизацией слоя гладких мышц сосудов, и SPA – VSMCs постепенно исчезают по мере продвижения процесса ремоделирования. В конце концов, происходит полная потеря мышечно-эластической артериальной среды ремоделированных артерий [4].
Исследование продемонстрировало вакуолизацию эндотелиальных клеток и набухание некоторых отдельных SPA гладкомышечных клеток сосудов (SPA – VSMC) на очень ранней стадии процесса ремоделирования [4].Инвазия EVT связана с дальнейшей дезорганизацией слоя гладких мышц сосудов, и SPA – VSMCs постепенно исчезают по мере продвижения процесса ремоделирования. В конце концов, происходит полная потеря мышечно-эластической артериальной среды ремоделированных артерий [4].
Ранние исследования продемонстрировали апоптоз SPA-VSMCs во время процесса ремоделирования, и было показано, что децидуальные иммунные клетки играют причинный эффект [5-7]. Этот момент ставится под сомнение находками, показывающими сигналы апоптоза преимущественно в EVT, децидуальных лимфоцитах и эндотелиальных клетках сосудов, но мало в SPA-VSMC [8].Исследование также подтвердило миграцию SPA – VSMCs в строму децидуальной оболочки после ремоделирования SPA [8]. Тем не менее, SPA – VSMC практически не отслеживаются в децидуальной оболочке с использованием традиционных маркеров VSMC.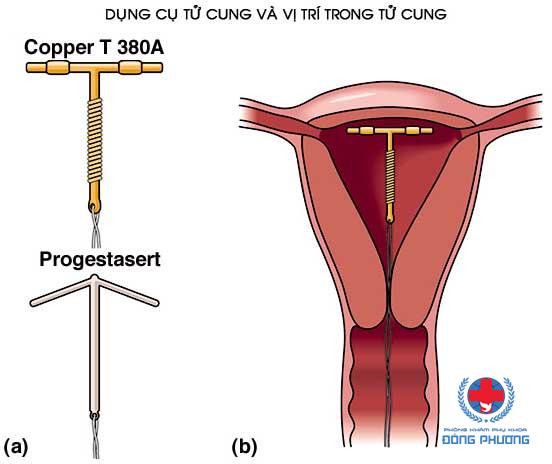 В последние годы появилось больше свидетельств, указывающих на дедифференцировку SPA-VSMC, о чем свидетельствует сниженная экспрессия типичных маркеров VSMC, таких как альфа-актин гладких мышц (αSMA), белок гладких мышц 22-альфа (SM22α), кальпонин, тяжелая цепь миозина. 11 ( MYh21 ), морфологическое преобразование в округлое, а также смещение или изменение картины распределения во время ремоделирования сосудов [9–11].Однако убедительные доказательства участия динамической дедифференцировки SPA – VSMCs во время реконструкции сосудов матки остаются недостаточными. Регуляторные механизмы дедифференцировки SPA-VSMCs человека и конечная судьба SPA-VSMCs остаются загадочными.
В последние годы появилось больше свидетельств, указывающих на дедифференцировку SPA-VSMC, о чем свидетельствует сниженная экспрессия типичных маркеров VSMC, таких как альфа-актин гладких мышц (αSMA), белок гладких мышц 22-альфа (SM22α), кальпонин, тяжелая цепь миозина. 11 ( MYh21 ), морфологическое преобразование в округлое, а также смещение или изменение картины распределения во время ремоделирования сосудов [9–11].Однако убедительные доказательства участия динамической дедифференцировки SPA – VSMCs во время реконструкции сосудов матки остаются недостаточными. Регуляторные механизмы дедифференцировки SPA-VSMCs человека и конечная судьба SPA-VSMCs остаются загадочными.
Ниша эндометрия матки, в которой присутствуют SPA – VSMC, претерпевает огромные изменения во время беременности. Децидуализация стромальных клеток матки начинается в поздне-лютеиновой фазе менструального цикла и прогрессирует очень выраженно и охватывает все отделы матки на протяжении беременности, с заметно повышенной секреторной способностью рекрутирования децидуальных клеток или размножения материнских лимфоцитов, преимущественно децидуальных естественных клеток-киллеров (dNKs). , и макрофаги, в децидуальный компартмент происходит инвазия EVT [12].Были проведены исследования, указывающие на ключевую роль децидуальных лимфоцитов и EVT в ремоделировании сосудов матки [7, 13, 14]. Дальнейшие исследования необходимы для выяснения динамически скоординированного эффекта нескольких типов клеток в децидуальной нише для модуляции клеточных свойств VSMC во время ремоделирования сосудов.
, и макрофаги, в децидуальный компартмент происходит инвазия EVT [12].Были проведены исследования, указывающие на ключевую роль децидуальных лимфоцитов и EVT в ремоделировании сосудов матки [7, 13, 14]. Дальнейшие исследования необходимы для выяснения динамически скоординированного эффекта нескольких типов клеток в децидуальной нише для модуляции клеточных свойств VSMC во время ремоделирования сосудов.
В настоящем исследовании, чтобы получить целостное представление о трансформации SPA – VSMC, мы проанализировали клеточные особенности SPA – VSMCs на разных стадиях ремоделирования SPA у человека на ранних сроках беременности. In vitro эксперименты на линии клеток VSMC были проведены для изучения влияния децидуальных стромальных клеток (DSC), dNKs, децидуальных макрофагов и EVT на дедифференцировку SPA – VSMC. Важно отметить, что потенциальная трансформация VSMC в CD56-позитивные dNKs была предположена с помощью иммунофлуоресценции-ДНК гибридизации-лигирования in-situ (IF-ISH-PLA) и анализа ChIP для модификации h4K4dime в промоторной области MYh21 .
Материалы и методы
Сбор проб
Нормальные децидуальные ткани человека на 5–12 неделях гестации были собраны после терапевтического прерывания здоровой беременности в больнице 306 PLA (Пекин, Китай).Ткани погружали в ледяную среду RPMI-1640 и подвергали первичной изоляции клеток или фиксации ткани в течение 1 часа после операции. Сбор человеческих образцов был разрешен местными этическими комитетами Института зоологии Китайской академии наук.
Выделение клеток и культивирование клеток
Первичные децидуальные клетки были изолированы из свежих децидуальных тканей в соответствии с предыдущими сообщениями [15–18]. Ткани децидуальной оболочки человека разрезали на небольшие кусочки размером 1 мм 3 с помощью GentelMACS Dissociator (Miltenyi Biotec, 130-093-235, Германия) и расщепляли 10 ед / мл ДНКазой типа I (DN25, Sigma Aldrich, St. Луис, Миссури, США) и 1,0 мг / мл коллагеназы типа IV (Gibco,
Луис, Миссури, США) и 1,0 мг / мл коллагеназы типа IV (Gibco,
Для выделения первичных ДСК [15] расщепленные децидуальные клетки последовательно фильтровали через ячейки 100, 70 и 40 мкм и осаждали центрифугированием при 800 × g в течение 10 мин. Осадок клеток ресуспендировали в DMEM (Hyclone, Sh40022.01B), добавляли 10% FBS (Gibco, 10270-106) и культивировали при 37 ° C в течение 30 минут для удаления быстро прикрепляющихся эпителиальных клеток.После дополнительной ночной культуры плавающие клетки удаляли, а прикрепленные клетки представляли собой клетки DSC.
Для выделения dNK [16] расщепленные децидуальные клетки культивировали в течение ночи в RPMI-1640 (Gibco, 31800-022) с добавлением 10% FBS (Gibco, 10270-106), и плавающие клетки подвергали очистке. dNK с помощью магнитных шариков, меченных антителом против CD56 (Miltenyi, 130050401) в соответствии с инструкциями производителя.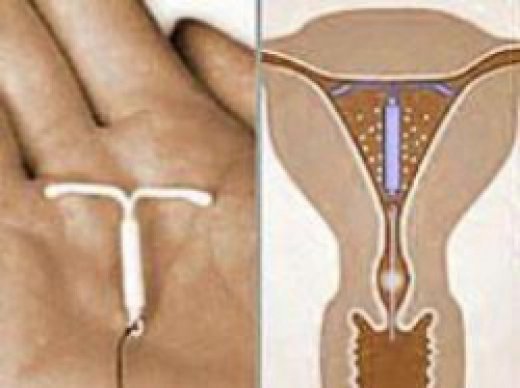 Вкратце, CD56-магнитные шарики добавляли к плавающим клеткам и инкубировали в течение 30 минут при 4 ° C с последующим сбором клеток с помощью колонок LS (Miltenyi, 130042401).Очищенные dNK культивировали в среде RPMI-1640 с добавлением 10% FBS при 37 ° C.
Вкратце, CD56-магнитные шарики добавляли к плавающим клеткам и инкубировали в течение 30 минут при 4 ° C с последующим сбором клеток с помощью колонок LS (Miltenyi, 130042401).Очищенные dNK культивировали в среде RPMI-1640 с добавлением 10% FBS при 37 ° C.
Для выделения децидуальных макрофагов [17] расщепленные децидуальные клетки обрабатывали магнитными шариками, меченными антителом к CD14 (Miltenyi, 130050201), в соответствии с инструкциями производителя. Вкратце, к клеткам добавляли магнитные шарики CD14 и инкубировали в течение 30 мин при 4 ° C с последующим сбором клеток с помощью колонок LS (Miltenyi, 130042401). Очищенные децидуальные клетки макрофагов культивировали в RPMI-1640 с добавлением 10% FBS при 37 ° C.
Изоляция EVT была выполнена в соответствии с предыдущим отчетом [18]. Расщепленные децидуальные клетки культивировали в течение ночи в RPMI-1640 (Gibco, 31800-022) с добавлением 10% FBS (Gibco, 10270-106), а плавающие клетки отбрасывали. Прилипшие клетки подвергали сортировке в потоке с помощью PE-меченых антител против HLA-G (Sigma Aldrich, SAB4700317, RRID: AB_10898912). Очищенные EVT культивировали в RPMI-1640 с добавлением 10% FBS при 37 ° C.
Очищенные EVT культивировали в RPMI-1640 с добавлением 10% FBS при 37 ° C.
Кондиционированные среды из DSC (DSC-CM) получали через 48 часов культивирования, а кондиционированные среды из dNK (dNK-CM), децидуальных макрофагов (dM-CM) и EVT (EVT-CM) получали отдельно через 24 часа. ч культуры клеток.Культуральные среды, свободные от клеток, собирали параллельно в качестве соответствующих контрольных сред (DSC-CTRL, dNK-CTRL, dM-CTRL и EVT-CTRL). Собранные среды хранили в стерильных пробирках и центрифугировали при 800 × g в течение 10 мин, а супернатанты хранили при -80 ° C и использовали в течение 1 месяца.
Клеточная линия T / G HA-VSMC (CRL-1999) была приобретена в ATCC и культивирована в среде F-12 K (Gibco, 21127022) с добавлением 10% FBS (Gibco, 10270106) и 0,01 мг / мл инсулина. , 0,01 мг / мл трансферрина, 10 нг / мл селенита натрия (Sigma, I1884), 0.05 мг / мл аскорбиновой кислоты (Sigma, A4403) и 0,03 мг / мл добавки для роста эндотелиальных клеток (Sigma, B211-GS). Клетки пассировали в соотношении 1: 3 каждые 3 дня. Для эксперимента по обработке условий среды клетки T / G HA-VSMC высевали в 6-луночный планшет из расчета 2 × 10 5 клеток / лунку, и добавляли различные среды для условий при 50% концентрации. Клетки обрабатывали в течение 3 дней и подвергали иммунофлуоресцентному анализу, проточной цитометрии или измерению различных молекул, как описано ниже.
Клетки пассировали в соотношении 1: 3 каждые 3 дня. Для эксперимента по обработке условий среды клетки T / G HA-VSMC высевали в 6-луночный планшет из расчета 2 × 10 5 клеток / лунку, и добавляли различные среды для условий при 50% концентрации. Клетки обрабатывали в течение 3 дней и подвергали иммунофлуоресцентному анализу, проточной цитометрии или измерению различных молекул, как описано ниже.
Чтобы вызвать апоптоз клеток, клетки T / G HA-VSMC обрабатывали 2 мкг / мл пуромицина (Solarbio, P8230, США) в течение 24 часов и подвергали измерению апоптоза клеток, как описано ниже.
Иммунофлуоресценция
Децидуальные ткани человека были зафиксированы и встроены в O.C.T. компаунда (Sakura Finetek, Торранс, Калифорния, США) и разрезали на 10 мкм. Замороженные срезы или культивированные клетки T / G HA-VSMC ненадолго фиксировали в 4% параформальдегиде и обрабатывали 0.1% тритона и подвергали инкубации со специфическими антителами против цитокератина 7 (CK7, ZSGB-BIO, P7000, Beijing, ZM-0071, 1: 200, RRID: AB_2864330), SM22α (Abcam, ab10135, 1: 200, RRID : AB_2255631, Кембридж, США), MYh21 (Abcam, ab53219, 1: 500, RRID: AB_2147146), кальпонин (Abcam, ab46794, 1: 200, RRID: AB_2147146), CD31 (Abcam, ab28364, 1: 100, RRID: AB_726362), αSMA (Millipore, CBL-171, 1: 400, RRID: AB_2223166, Массачусетс, США) или CD56 (Abcam, ab75813, 1: 300 RRID: AB_2632384).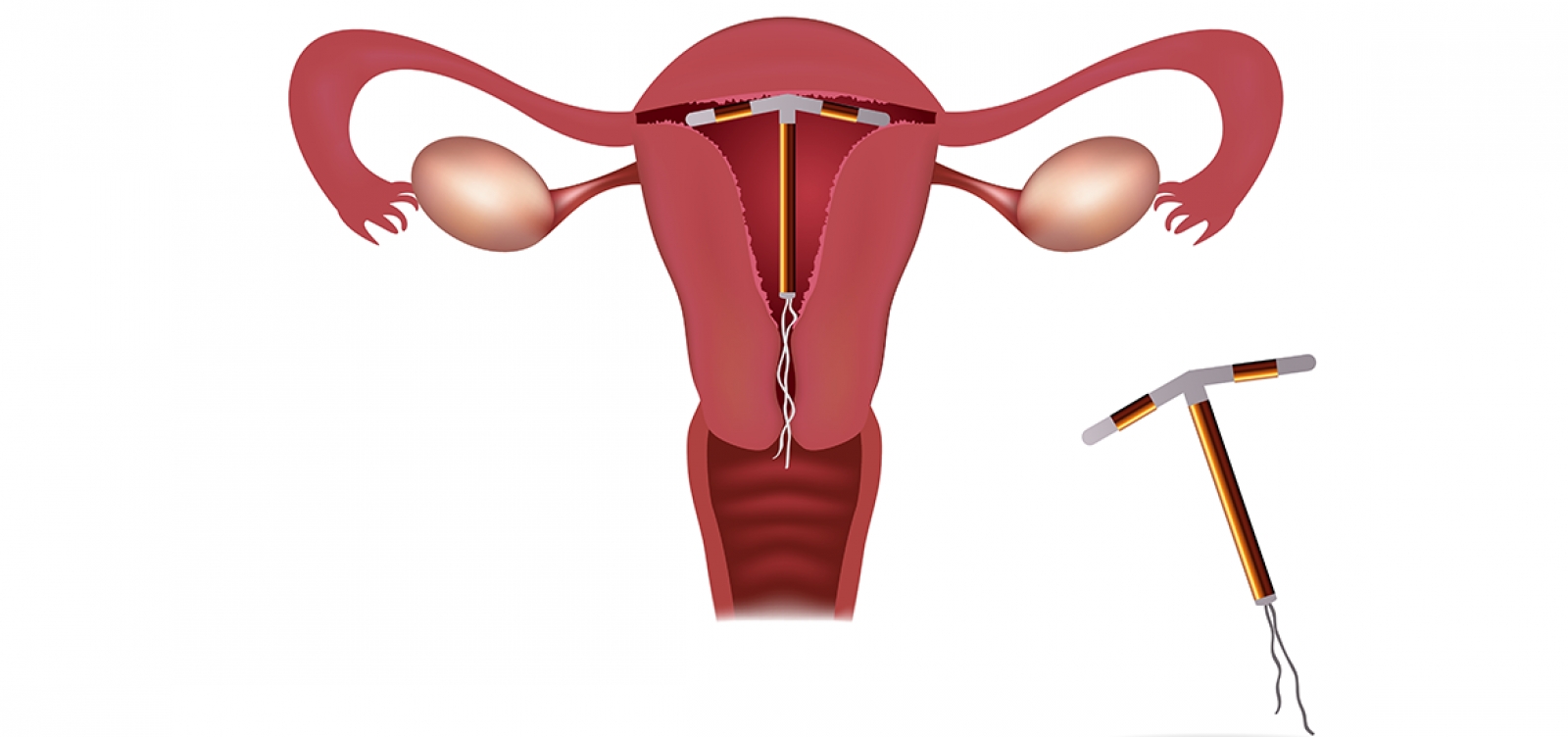 Антитела визуализировали с использованием вторичных антител, конъюгированных с FITC или TRITC (ZSGB-BIO, ZF-0311, ZF-0313, 1: 100, RRID: AB_2571576, RRID: AB_2571577). Ядра клеток окрашивали 4,6-диамидино-2-фенилиндолом (DAPI; Sigma, 28718-90-3). Результаты регистрировались конфокальной системой Zeiss LSM780 (Zeiss, Oberkochen, BW, Германия) и обрабатывались с помощью программного обеспечения ZEN 2012 (Zeiss).
Антитела визуализировали с использованием вторичных антител, конъюгированных с FITC или TRITC (ZSGB-BIO, ZF-0311, ZF-0313, 1: 100, RRID: AB_2571576, RRID: AB_2571577). Ядра клеток окрашивали 4,6-диамидино-2-фенилиндолом (DAPI; Sigma, 28718-90-3). Результаты регистрировались конфокальной системой Zeiss LSM780 (Zeiss, Oberkochen, BW, Германия) и обрабатывались с помощью программного обеспечения ZEN 2012 (Zeiss).
Иммуногистохимия
Децидуальные ткани человека фиксировали в 4% параформальдегиде и подвергали обычной дегидратации и заливке парафином.Парафиновые срезы толщиной 5 мкм подвергали стандартной регидратации, извлечению антигена и блокированию перед инкубацией со специфическим антителом против CK7 (ZSGB-BIO, ZM-0071, 1: 200, RRID: AB_2864330), αSMA (Abcam, ab5694 , 1: 1000, RRID: AB_2223021) и Виментин (ZSGB-BIO, ZM-0260, 1: 100, RRID: AB_10562134). Выделенные первичные ДСК фиксировали в 4% параформальдегиде, обрабатывали 0,1% тритоном и подвергали инкубации со специфическими антителами против виментина (Abcam, ab
, 1: 500, RRID: AB_10562134). Срезы или клетки дополнительно инкубировали со вторыми антителами, конъюгированными с HRP (ZSGB-BIO, PV-6001, PV-6002, RRID: AB_2864333, RRID: AB_2864334, Пекин, Китай) и DAB (ZSGB-BIO, ZLI-9019 ) субстрата с последующим контрастным окрашиванием гематоксилином и закреплением. Изображения регистрировали на световом микроскопе с ПЗС-матрицей (DP72, Olympus, Япония).
Срезы или клетки дополнительно инкубировали со вторыми антителами, конъюгированными с HRP (ZSGB-BIO, PV-6001, PV-6002, RRID: AB_2864333, RRID: AB_2864334, Пекин, Китай) и DAB (ZSGB-BIO, ZLI-9019 ) субстрата с последующим контрастным окрашиванием гематоксилином и закреплением. Изображения регистрировали на световом микроскопе с ПЗС-матрицей (DP72, Olympus, Япония).Подготовка РНК и количественная ПЦР в реальном времени
Суммарную РНК из культивированных T / G HA-VSMC экстрагировали реагентом TRIzol в соответствии с инструкциями производителя (Invitrogen, 155, Carlsbad, CA, USA).Обратную транскрипцию проводили с использованием Oligo Primer (TIANGEN, CD106, Пекин, Китай), а кДНК использовали в качестве матрицы для количественной ПЦР в реальном времени с помощью набора SYBR Premix Ex TaqTM (Takara, DRR820A, Далянь, Китай) в соответствии с инструкции производителя. Последовательности праймеров показаны в дополнительной таблице S1.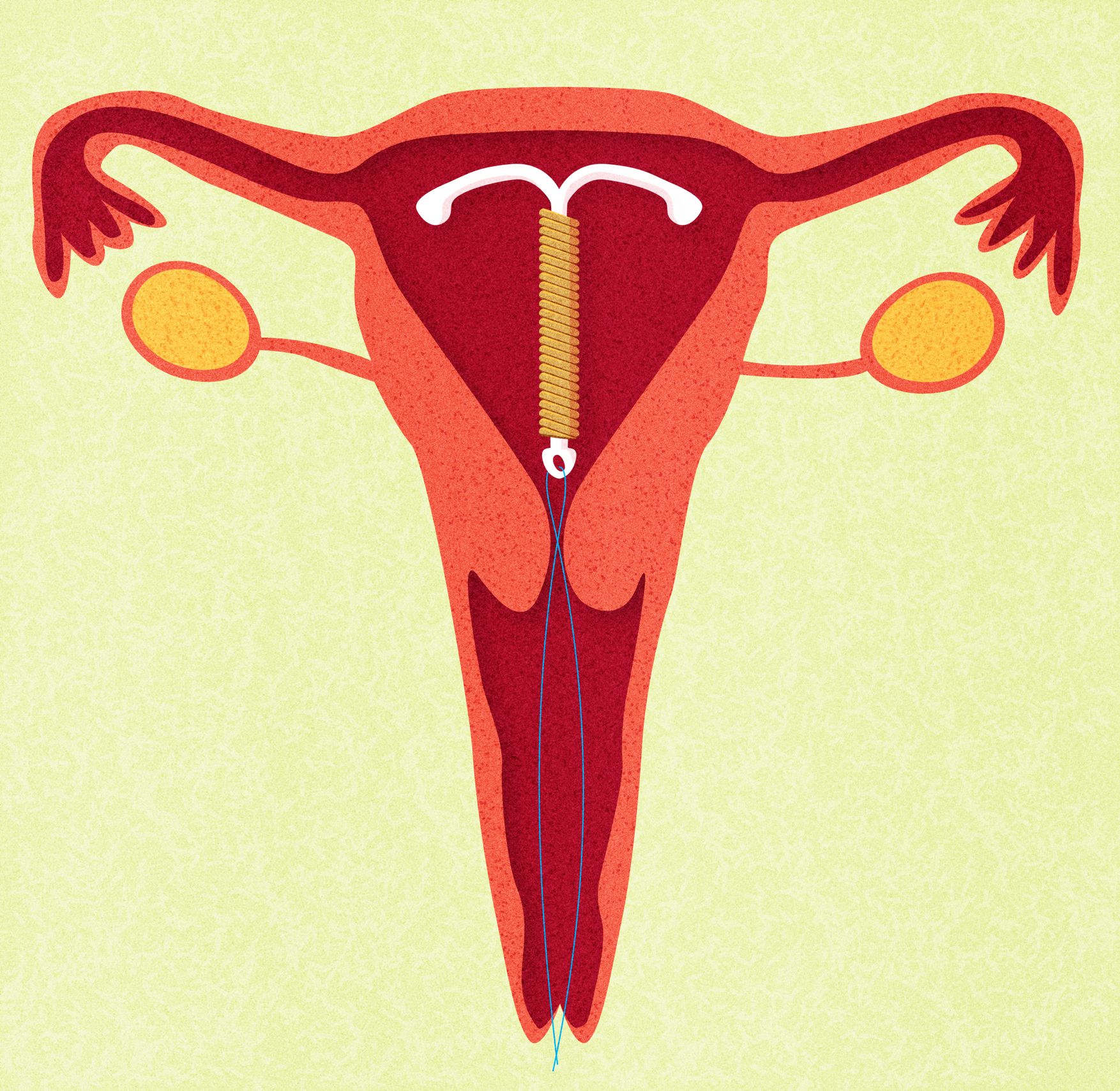 После процесса реакции ПЦР относительную экспрессию определенного гена нормализовали с помощью Gapdh с использованием метода 2 -ΔΔCT .
После процесса реакции ПЦР относительную экспрессию определенного гена нормализовали с помощью Gapdh с использованием метода 2 -ΔΔCT .
Приготовление белка и вестерн-блоттинг
Общий белок T / G HA-VSMC экстрагировали буфером для лизиса RIPA и 10 мкг белка подвергали 10% SDS-PAGE и последующему электропереносу на нитроцеллюлозную мембрану (Amersham Pharmacia Biotech, Бакингемшир, Великобритания).Антитела, использованные в этом исследовании, включали αSMA (Abcam, ab5694, 1: 1000, RRID: AB_2223021), SM22α (Abcam, ab10135, 1: 10000, RRID: AB_2255631), кальпонин (Abcam, ab46794, 1: 10000, RRID: AB_22
) , расщепленная каспаза3 (Cell Signaling Technology, 9661, 1: 1000, RRID: AB_2341188, Массачусетс, США) и тубулин (Abcam, ab7291, 1: 10000, RRID: AB_2241126). Вторичные антитела, конъюгированные с HRP (W4021, Promega, 1: 5000, RRID: AB_430834; W4011, Promega, 1: 5000, RRID: AB_430833, Висконсин, США; SA00001-4, Proteintech Group, 1: 5000, RRID: AB_2864335, Чикаго , США) использовали в соответствии с видами первичных антител.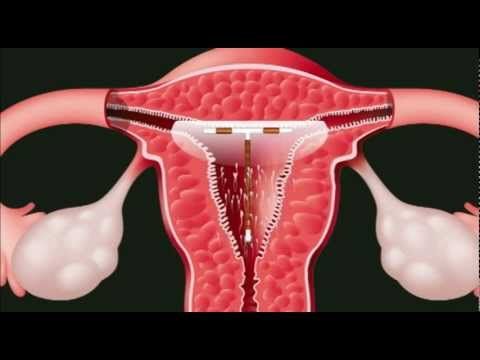 Для визуализации применялась система усиленной хемилюминесценции (Thermo Scientific, 34580, Массачусетс, США). Относительную плотность обнаруженной молекулы нормализовали по значению тубулина в том же блоте.
Для визуализации применялась система усиленной хемилюминесценции (Thermo Scientific, 34580, Массачусетс, США). Относительную плотность обнаруженной молекулы нормализовали по значению тубулина в том же блоте.Анализ пролиферации клеток
Cell Counting Kit-8 (CCK-8) (Dojindo, CK04, Япония) использовали в соответствии с инструкциями производителя. Вкратце, 10 мкл раствора CCK-8 добавляли в каждую лунку для культивирования и после культивирования в течение 4 ч определяли оптическую плотность при 450 нм с помощью считывающего устройства для микропланшетов.
Рисунок 1
Динамическая трансформация SPA – VSMCs в процессе ремоделирования SPA в децидуальной оболочке беременных на ранних сроках. (A) Иммунофлуоресцентное окрашивание CD31 (зеленый) и CK7 (красный), αSMA (зеленый) в соседних срезах для идентификации различных стадий ремоделирования SPA. (B) Иммунофлуоресцентное окрашивание CD31 (зеленый) и CK7 (красный), αSMA (зеленый) и SM22α (красный), кальпонина (красный) в соседних срезах SPA на разных стадиях ремоделирования.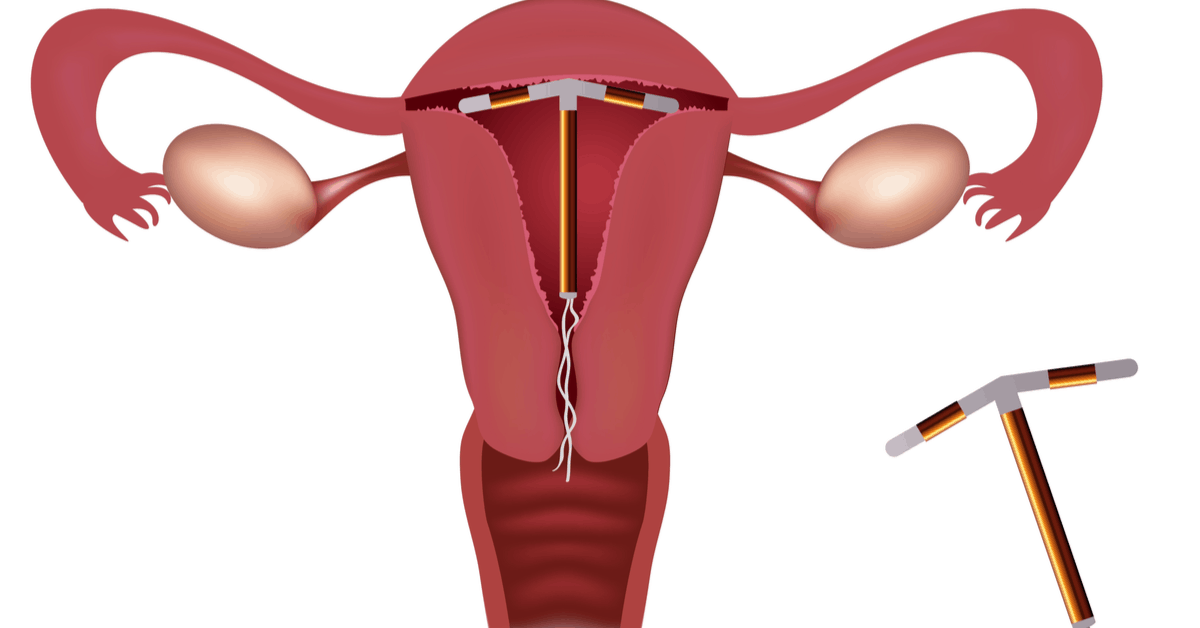 Увеличение прямоугольных областей, показывающих закругленные, разделенные и смещенные SPA – VSMC.Розовые стрелки указывают закругленные SPA-VSMC, желтые стрелки указывают разделенные SPA-VSMC, а белые стрелки указывают смещенные SPA-VSMC. (C) Статистический анализ морфологически трансформированных SPA – VSMCs на разных стадиях ремоделирования. Левая панель представляет собой частоту разделенных SPA, а правая панель показывает долю округленных и смещенных SPA – VSMC. Статистический анализ выполняется на основе записи из пяти случаев для каждого этапа ремоделирования и трех случайных просмотров в каждом случае.Данные представлены в виде среднего значения ± стандартное отклонение, сравнение между группами проводится с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Увеличение прямоугольных областей, показывающих закругленные, разделенные и смещенные SPA – VSMC.Розовые стрелки указывают закругленные SPA-VSMC, желтые стрелки указывают разделенные SPA-VSMC, а белые стрелки указывают смещенные SPA-VSMC. (C) Статистический анализ морфологически трансформированных SPA – VSMCs на разных стадиях ремоделирования. Левая панель представляет собой частоту разделенных SPA, а правая панель показывает долю округленных и смещенных SPA – VSMC. Статистический анализ выполняется на основе записи из пяти случаев для каждого этапа ремоделирования и трех случайных просмотров в каждом случае.Данные представлены в виде среднего значения ± стандартное отклонение, сравнение между группами проводится с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 1
Динамическое преобразование SPA – VSMCs в процессе ремоделирования SPA в децидуальной оболочке беременных на ранних сроках. (A) Иммунофлуоресцентное окрашивание CD31 (зеленый) и CK7 (красный), αSMA (зеленый) в соседних срезах для идентификации различных стадий ремоделирования SPA. (B) Иммунофлуоресцентное окрашивание CD31 (зеленый) и CK7 (красный), αSMA (зеленый) и SM22α (красный), кальпонина (красный) в соседних срезах SPA на разных стадиях ремоделирования.Увеличение прямоугольных областей, показывающих закругленные, разделенные и смещенные SPA – VSMC. Розовые стрелки указывают закругленные SPA-VSMC, желтые стрелки указывают разделенные SPA-VSMC, а белые стрелки указывают смещенные SPA-VSMC. (C) Статистический анализ морфологически трансформированных SPA – VSMCs на разных стадиях ремоделирования. Левая панель представляет собой частоту разделенных SPA, а правая панель показывает долю округленных и смещенных SPA – VSMC. Статистический анализ выполняется на основе записи из пяти случаев для каждого этапа ремоделирования и трех случайных просмотров в каждом случае.Данные представлены в виде среднего значения ± стандартное отклонение, сравнение между группами проводится с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
(B) Иммунофлуоресцентное окрашивание CD31 (зеленый) и CK7 (красный), αSMA (зеленый) и SM22α (красный), кальпонина (красный) в соседних срезах SPA на разных стадиях ремоделирования.Увеличение прямоугольных областей, показывающих закругленные, разделенные и смещенные SPA – VSMC. Розовые стрелки указывают закругленные SPA-VSMC, желтые стрелки указывают разделенные SPA-VSMC, а белые стрелки указывают смещенные SPA-VSMC. (C) Статистический анализ морфологически трансформированных SPA – VSMCs на разных стадиях ремоделирования. Левая панель представляет собой частоту разделенных SPA, а правая панель показывает долю округленных и смещенных SPA – VSMC. Статистический анализ выполняется на основе записи из пяти случаев для каждого этапа ремоделирования и трех случайных просмотров в каждом случае.Данные представлены в виде среднего значения ± стандартное отклонение, сравнение между группами проводится с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.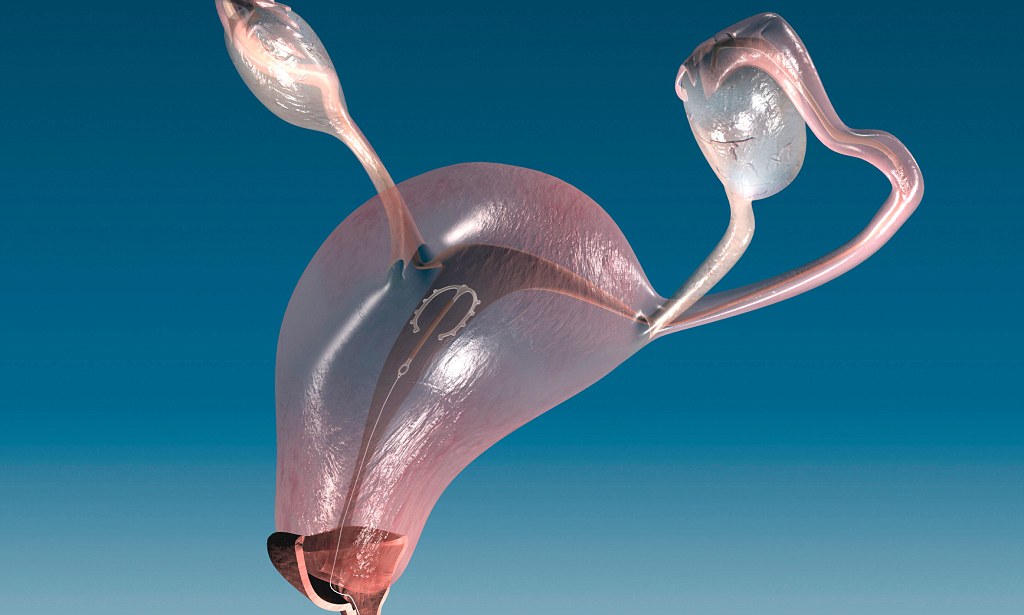
Иммунофлуоресценция – ДНК – ISH – PLA
Эксперимент проводился, как сообщалось ранее [19]. Вкратце, меченный биотином зонд MYh21 человека был получен с помощью системы маркировки ДНК BioNick (Thermo Fisher, 18247015, США). Последовательность праймера приведена в дополнительной таблице S1.Иммунофлуоресценцию для CK7, виментина, αSMA и CD56 проводили в парафиновом срезе децидуальных тканей, как описано выше, и срезы переваривали пепсином (ZSGB-BIO) при 37 ° C в течение 10 мин. Зонд MYh21 денатурировали в течение 5 мин при 80 ° C и наносили на срезы. Срезы инкубировали при 37 ° C в течение 16 ч с последующей промывкой буфером SSC и дальнейшей инкубацией с антителами против h4K4dime (05-1338, Millipore, RRID: AB_1977248) и биотина (Abcam, ab53494, RRID: AB_867860) в течение ночи при 4. ° C.PLA выполняли с использованием набора для обнаружения Duolink Orange и наблюдали при 555 нм (DUO
, DUO
, DUO, Sigma, США) в соответствии с инструкциями производителя. Ядра клеток окрашивали 4,6-диамидино-2-фенилиндолом (DAPI; Sigma, 28718-90-3). Результаты регистрировались конфокальной системой Zeiss LSM780 (Zeiss, Oberkochen, BW, Германия) и обрабатывались с помощью программного обеспечения ZEN 2012 (Zeiss).
Ядра клеток окрашивали 4,6-диамидино-2-фенилиндолом (DAPI; Sigma, 28718-90-3). Результаты регистрировались конфокальной системой Zeiss LSM780 (Zeiss, Oberkochen, BW, Германия) и обрабатывались с помощью программного обеспечения ZEN 2012 (Zeiss).Анализ ChIP
Выделенные EVT, DSC и dNK подвергали выделению хроматина с использованием набора Millipore EZ ChIP Kit (Millipore, 17-371, RRID: AB_1977248) в соответствии с инструкциями производителя.Вкратце, клетки были зафиксированы и сшиты 1% формальдегидом и остановлены раствором глицина. Затем клетки лизировали буфером для лизиса SDS и обрабатывали ультразвуком в течение 15 мин (9 с пробег и 30 с пауза на цикл) для получения фрагментов ДНК длиной 200–2000 п.н. Разрезанные фрагменты ДНК связывали с белком G агарозы в течение 1 часа и инкубировали с антителом h4K4dime (Millipore, 05-1338) при 4 ° C в течение ночи. После промывки использовали буфер для элюции и 5 M Nacl для высвобождения поперечной сшивки, а РНКазу A использовали для гидролиза загрязненной РНК. ДНК очищали и анализировали с помощью количественной ПЦР в реальном времени. Последовательности праймеров перечислены в дополнительной таблице S1.
ДНК очищали и анализировали с помощью количественной ПЦР в реальном времени. Последовательности праймеров перечислены в дополнительной таблице S1.
Проточная цитометрия
Для анализа чистоты клеток выделенные dNK, децидуальные макрофаги и EVT отдельно инкубировали со следующими антителами в течение 30 мин при 4 ° C: меченные APC анти-CD56 (BioLegend, 318310, RRID: AB_604106, San Diego CA, США), меченный FITC анти-CD14 (BioLegend, 367115, RRID: AB_2571928) и меченный PE анти-HLA-G (ebioscience, 12-9957-42, RRID: AB_11149313, Калифорния, США) и подвергали обработке цитометрический анализ.
Рисунок 2
Локализация DSC, dNK, децидуальных макрофагов и EVT на разных стадиях ремоделирования SPA. (A) Типичный результат иммуногистохимии для CK7, αSMA и виментина в ткани децидуальной оболочки с артериями Un-Rem на 5 неделе гестации. (B) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD56 (белый) и CK7 (красный) во время СПА-ремоделирование децидуальной оболочки на ранних сроках беременности.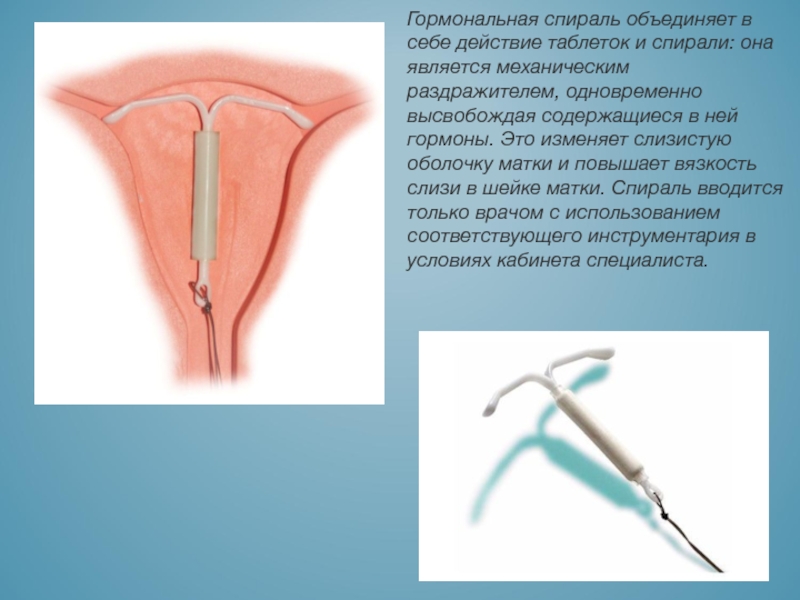 Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (C) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD163 (белый) и CK7 (красный) во время ремоделирования SPA в децидуальной оболочке беременных на ранних сроках.Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (D) Иммунофлуоресцентное окрашивание CK7 (красный) и SM22α (зеленый) во время ремоделирования SPA у децидуальной оболочки на ранних сроках беременности. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. Стрелки указывают смещенные и разделенные SPA – VSMC. Шкала показывает 100 мкм.
Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (C) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD163 (белый) и CK7 (красный) во время ремоделирования SPA в децидуальной оболочке беременных на ранних сроках.Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (D) Иммунофлуоресцентное окрашивание CK7 (красный) и SM22α (зеленый) во время ремоделирования SPA у децидуальной оболочки на ранних сроках беременности. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. Стрелки указывают смещенные и разделенные SPA – VSMC. Шкала показывает 100 мкм.
Рисунок 2
Локализация DSC, dNK, децидуальных макрофагов и EVT на разных стадиях ремоделирования SPA. (A) Типичный результат иммуногистохимии для CK7, αSMA и виментина в ткани децидуальной оболочки с артериями Un-Rem на 5 неделе гестации.(B) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD56 (белый) и CK7 (красный) во время ремоделирования SPA у децидуальной оболочки беременных на ранних сроках.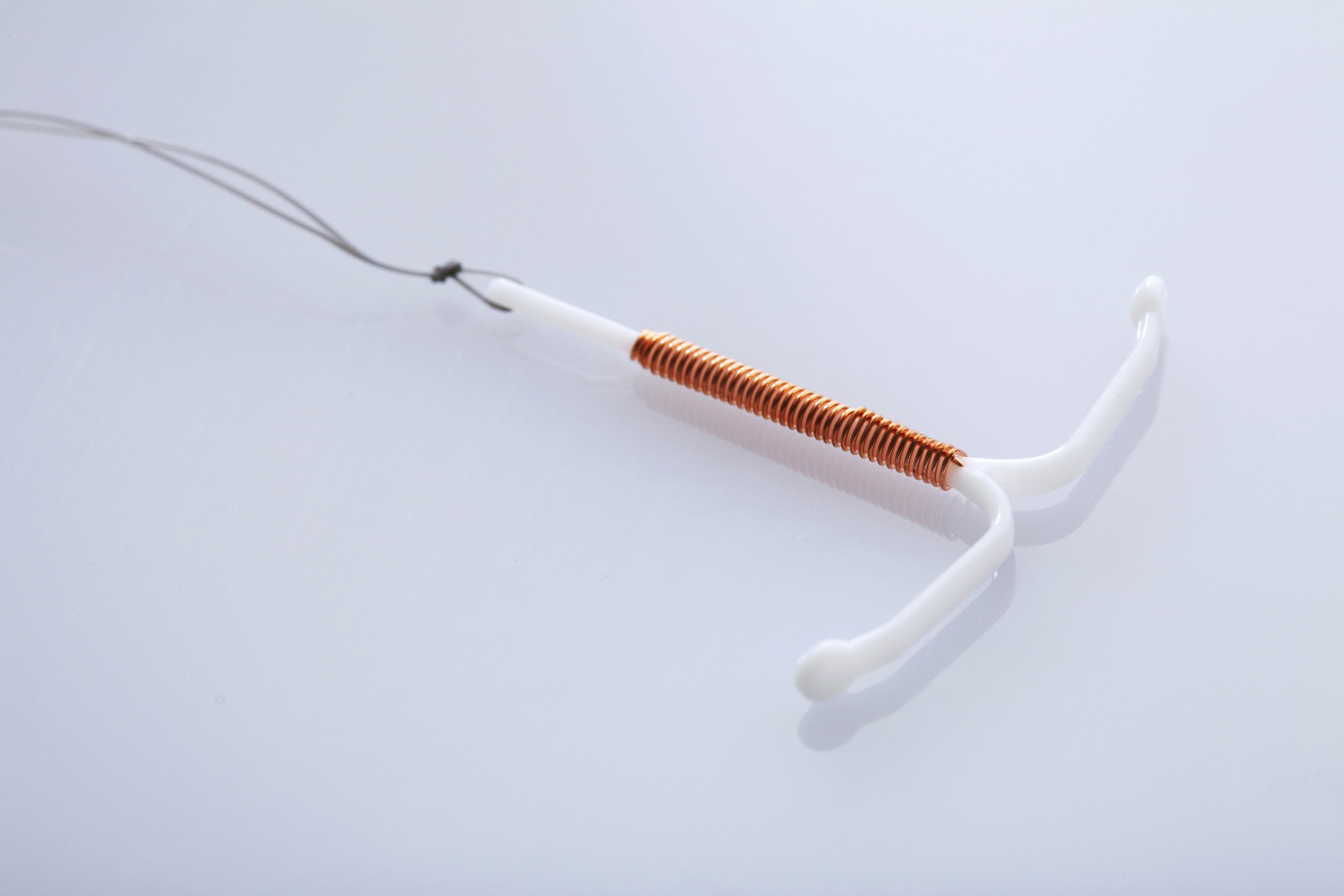 Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (C) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD163 (белый) и CK7 (красный) во время ремоделирования SPA в децидуальной оболочке беременных на ранних сроках. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (D) Иммунофлуоресцентное окрашивание CK7 (красный) и SM22α (зеленый) во время ремоделирования SPA у децидуальной оболочки на ранних сроках беременности. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA.Стрелки указывают смещенные и разделенные SPA – VSMC. Шкала показывает 100 мкм.
Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (C) Иммунофлуоресцентное окрашивание SM22α (зеленый), CD163 (белый) и CK7 (красный) во время ремоделирования SPA в децидуальной оболочке беременных на ранних сроках. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA. (D) Иммунофлуоресцентное окрашивание CK7 (красный) и SM22α (зеленый) во время ремоделирования SPA у децидуальной оболочки на ранних сроках беременности. Верхняя панель показывает Un-Rem SPA, а нижняя панель показывает реконструированную SPA.Стрелки указывают смещенные и разделенные SPA – VSMC. Шкала показывает 100 мкм.
Для клеточного апоптоза и анализа жизнеспособности клеток клетки T / G HA-VSMC, обработанные различными средами для кондиционирования или индуцированные 2 мкг / мл пуромицина, расщепляли 0,01% трипсином и подвергали воздействию аннексина V / PI (трансген, FA101 , Пекин, Китай) и 7-AAD (ebioscience, 420403) соответственно. Вкратце, для анализа аннексина V / PI к суспендированным клеткам добавляли аннексин V-FITC и PI в соотношении 1:20 и инкубировали в течение 15 минут при комнатной температуре с последующим анализом проточной цитометрии в течение 1 часа. Для анализа 7-AAD к суспендированным клеткам добавляли 7-AAD в концентрации 0,25 мкг на миллион клеток и инкубировали при 4 ° C в течение 10 мин перед анализом проточной цитометрии. Анализ проточной цитометрии проводили с использованием CytoFLEX (Beckman Coulter, Inc.), а данные анализировали с помощью CytExpert (Beckman Coulter, Inc.).
Для анализа 7-AAD к суспендированным клеткам добавляли 7-AAD в концентрации 0,25 мкг на миллион клеток и инкубировали при 4 ° C в течение 10 мин перед анализом проточной цитометрии. Анализ проточной цитометрии проводили с использованием CytoFLEX (Beckman Coulter, Inc.), а данные анализировали с помощью CytExpert (Beckman Coulter, Inc.).
Рисунок 3
DSC активно индуцируют дедифференцировку VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных DSC-CTRL или DSC-CM ( n = 3) .(C) Иммунофлуоресцентное окрашивание αSMA в T / G HA-VSMC, обработанном DSC-CTRL или DSC-CM. Статистический анализ проводится по не менее чем трем независимо повторяющимся экспериментам. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05.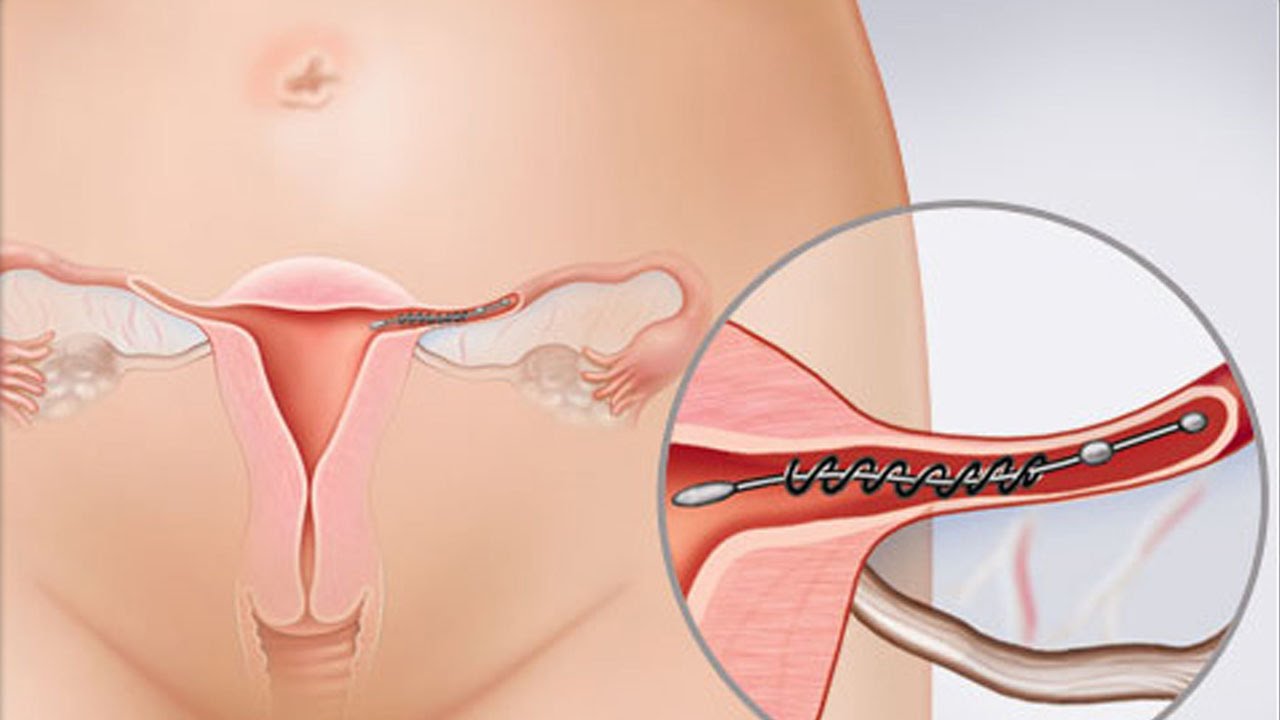 Шкала показывает 100 мкм.
Шкала показывает 100 мкм.
Рисунок 3
DSC активно индуцируют дедифференцировку VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных DSC-CTRL или DSC-CM ( n = 3) .(C) Иммунофлуоресцентное окрашивание αSMA в T / G HA-VSMC, обработанном DSC-CTRL или DSC-CM. Статистический анализ проводится по не менее чем трем независимо повторяющимся экспериментам. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Статистический анализ
Статистические анализы были выполнены с помощью GraphPad Prism (Сан-Диего, Калифорния, США) и представлены как среднее ± стандартное отклонение.Статистическое сравнение между группами выполняли с помощью независимого теста t или множественного сравнительного анализа с помощью теста Тьюки-Крамера в соответствии с по меньшей мере тремя независимо повторяемыми экспериментами.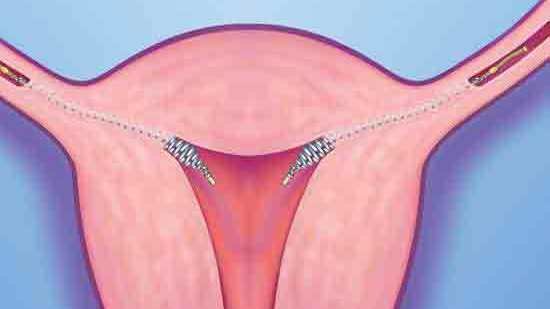 Значение P менее 0,05 считалось статистически значимым.
Значение P менее 0,05 считалось статистически значимым.
Результаты
Характеристики SPA – VSMC на разных этапах ремоделирования SPA человека
Децидуальные базальные клетки человека на 6–12 неделях гестации подвергали иммунофлуоресцентному анализу на CK7, CD31 и αSMA в соседних срезах, которые маркируют EVT, эндотелиальные клетки и VSMC соответственно.Основываясь на паттерне распределения этих клеток, SPA могут быть идентифицированы как Un-Rem, стадии раннего ремоделирования (Early-Rem), активного ремоделирования (Act-Rem) и стадии Fully-Rem (Figure 1A). Иммунное окрашивание на три традиционных маркера VSMC, αSMA, SM22α и кальпонин, было дополнительно выполнено для изучения морфологического изменения SPA – VSMC на этих стадиях ремоделирования SPA (рис. 1B). Интересно, что морфологические изменения SPA – VSMC, включая смещение и / или округлую форму, можно было обнаружить в Un-Rem SPA, хотя и с относительно низкой частотой. На стадии Early-Rem инвазивные EVT собирались вокруг SPA, и небольшое количество EVT можно было обнаружить на стенке сосуда. Многослойные VSMC вокруг этих SPA казались сильно ослабленными и разделенными, а многие VSMC выглядели значительно смещенными и / или круглыми. На стадии Act-Rem EVT в значительной степени заменяли эндотелий в SPA, в то время как количество VSMC было заметно уменьшено. В сосудах Fully-Rem окрашивание на αSMA, SM22α и кальпонин было неизменно неопределяемым (рис. 1B).Чтобы количественно оценить морфологические изменения SPA-VSMC, мы статистически проанализировали по крайней мере пять случаев для каждой стадии ремоделирования и записали три случайных изображения в каждом случае, чтобы подсчитать процент сосудов с разделенными слоями гладких мышц и VSMC с смещением и круглой формой. Как показано на Рисунке 1C, частота сосудов с разделением слоев и соотношение трансформированных VSMC были значительно выше на стадиях ремоделирования по сравнению со стадией Un-Rem, а изменения в артерии Act-Rem были более очевидными, чем в артерии Early-Rem.
На стадии Early-Rem инвазивные EVT собирались вокруг SPA, и небольшое количество EVT можно было обнаружить на стенке сосуда. Многослойные VSMC вокруг этих SPA казались сильно ослабленными и разделенными, а многие VSMC выглядели значительно смещенными и / или круглыми. На стадии Act-Rem EVT в значительной степени заменяли эндотелий в SPA, в то время как количество VSMC было заметно уменьшено. В сосудах Fully-Rem окрашивание на αSMA, SM22α и кальпонин было неизменно неопределяемым (рис. 1B).Чтобы количественно оценить морфологические изменения SPA-VSMC, мы статистически проанализировали по крайней мере пять случаев для каждой стадии ремоделирования и записали три случайных изображения в каждом случае, чтобы подсчитать процент сосудов с разделенными слоями гладких мышц и VSMC с смещением и круглой формой. Как показано на Рисунке 1C, частота сосудов с разделением слоев и соотношение трансформированных VSMC были значительно выше на стадиях ремоделирования по сравнению со стадией Un-Rem, а изменения в артерии Act-Rem были более очевидными, чем в артерии Early-Rem.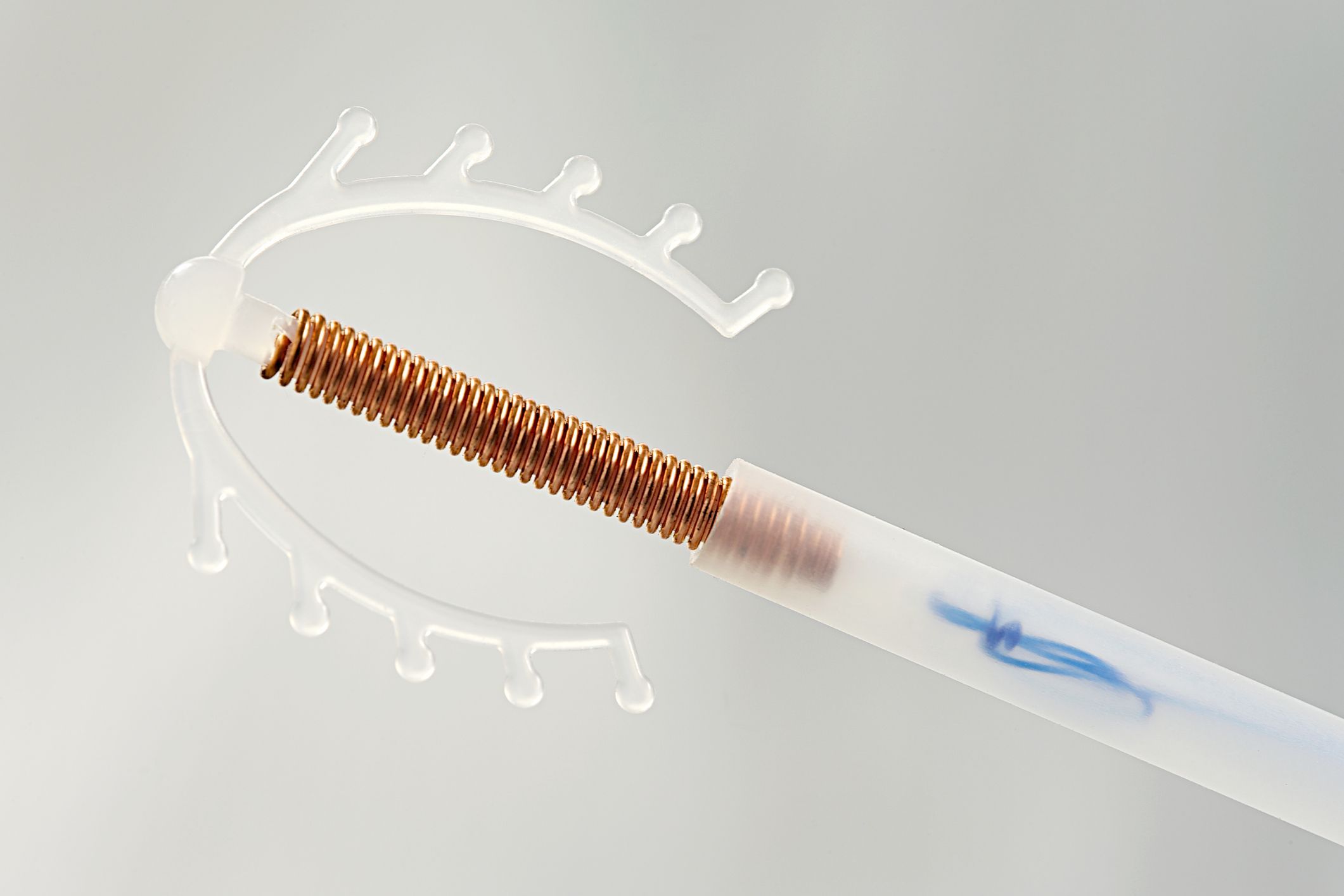 (Рисунок 1С).
(Рисунок 1С).
Взятые вместе, эти данные наблюдений четко иллюстрировали динамическую трансформацию SPA – VSMCs в процессе ремоделирования SPA.
Децидуальные СК индуцировали дедифференцировку VSMC
Результаты нашего гистологического исследования процесса ремоделирования сосудов матки подтверждают наличие трансформации VSMC до инвазии трофобластов. Литературные данные продемонстрировали, что SPA – VSMCs не проявляют трансформации в небеременном эндометрии [4, 20].При эмбриональной имплантации клетки стромы эндометрия претерпевают ярко выраженную децидуализацию, которая продуцирует множество цитокинов, хемокинов и гормонов и привлекает лимфоциты [21, 22]. Наш иммуногистохимический анализ на αSMA, виментин (маркировка эндометриальных SCs) и CK7 в соседних участках децидуальной основы человека на ранних сроках беременности выявил очень близкую локализацию DSC с смещенными SPA – VSMC вокруг артерий Un-Rem (рис. 2A). Таким образом, мы предположили, что клетки децидуальной стромы могут индуцировать трансформацию VSMCs на EVT-независимой стадии.
2A). Таким образом, мы предположили, что клетки децидуальной стромы могут индуцировать трансформацию VSMCs на EVT-независимой стадии.
Рисунок 4
Децидуальные NK способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных dNK-CTRL и dNK-CM ( n = 3 ). (C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с dNK-CTRL и dNK-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью теста Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 4
Децидуальные NK способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных dNK-CTRL и dNK-CM ( n = 3 ).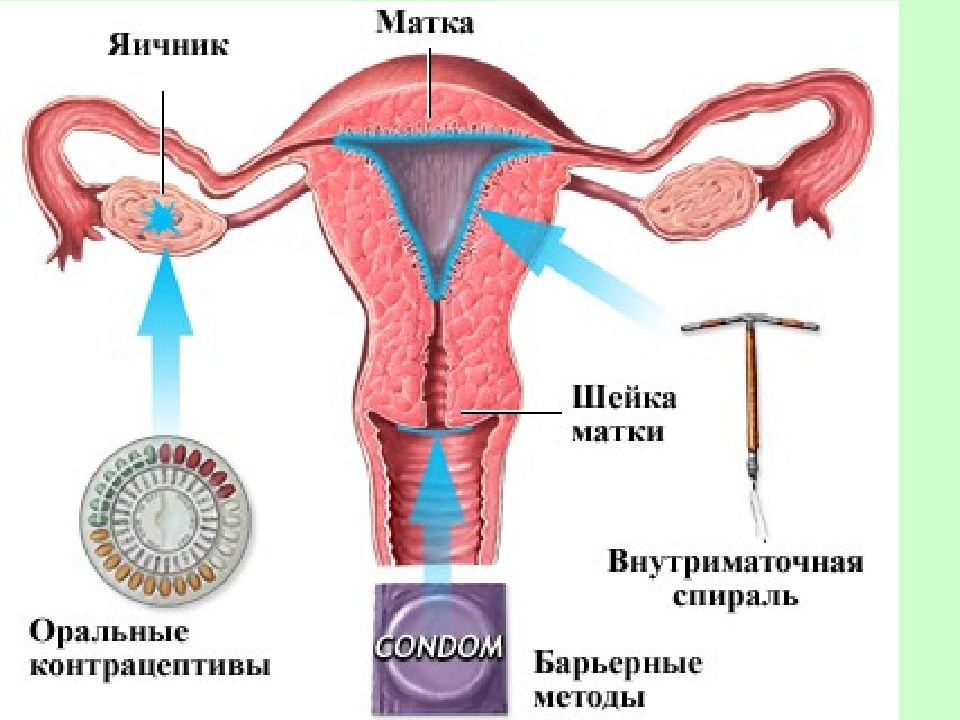 (C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с dNK-CTRL и dNK-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью теста Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
(C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с dNK-CTRL и dNK-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью теста Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 5
Децидуальные макрофаги способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных dM-CTRL и dM-CM ( n = 3 ). (C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с dM-CTRL и dM-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 5
Децидуальные макрофаги способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G HA-VSMC, обработанных dM-CTRL и dM-CM ( n = 3 ). (C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с dM-CTRL и dM-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Для измерения дедифференцировки VSMC в качестве модели in vitro использовали клеточную линию VSMC аорты человека, T / G HA-VSMC. Мы ферментативно выделили первичные DSC человека на ранних сроках беременности (дополнительный рисунок S1A) и собрали среду для условий после 48 часов культивирования (DSC-CM). Культуральные среды без DSC собирали параллельно в качестве контроля (DSC-CTRL). Клетки T / G HA-VSMC подвергали 3-дневной обработке DSC-CM или DSC-CTRL при 50% концентрации и измеряли экспрессию маркеров для SMC.Результаты ПЦР в реальном времени (рис. 3A) и вестерн-блоттинга (рис. 3B) показали, что уровни транскрипции и уровни белка αSMA, SM22α, кальпонина и MYh21 были значительно снижены в DSC-CM-обработанных клетках по сравнению с таковыми в клетках, обработанных DSC-CM. контрольные клетки, обработанные DSC-CTRL. Параллельно иммунофлуоресценция для αSMA показала очевидное уменьшение положительного сигнала αSMA и увеличение количества круглых клеток (рис. 3C).
Для оценки потенциального влияния лечения на жизнеспособность клеток, пролиферацию и апоптоз клеток анализировали в клетках T / G HA-VSMC после 3-дневной инкубации с кондиционирующей средой.Результат анализа CCK8 показал отсутствие изменений в росте клеток после обработки DSC-CM (дополнительная фигура S1E). Результаты проточной цитометрии для аннексина V / PI и 7-AAD, а также вестерн-блоттинга для расщепленной каспазы 3 продемонстрировали небольшую апоптотическую функцию в клетках T / G HA-VSMC, обработанных DSC-CTRL или DSC-CM, в то время как апоптоз в большом количестве наблюдался. сигналы в положительных контрольных клетках, которые были индуцированы пуромицином 2 мкг / мл (дополнительная фигура S2). Данные показали, что обработка DSC-CM в T / G HA-VSMC не влияла на жизнеспособность клеток.
Децидуальные NK-клетки и макрофаги способствовали дедифференцировке VSMC
Рекрутмент массивных лимфоцитов в децидуальную оболочку — критическое событие на ранних сроках беременности. Среди децидуальных лимфоцитов на ранних сроках беременности dNK и макрофаги составляют примерно 70 и 20% соответственно [23-25]. Эти клетки играют множество ролей, в том числе способствуют ремоделированию SPA, способствуют инвазии трофобластов и росту плода, регулируют дифференцировку других иммунных клеток и так далее [26–28].В образцах decidual basalis наш иммунофлуоресцентный анализ для SM22α, CD56 / CD163 и CK7 показал, что dNKs и макрофаги локализованы близко к трансформированным SPA – VSMC как на стадии Un-Rem, так и на стадии ремоделирования (рис. 2B и C). Эти лимфоциты потенциально могут способствовать дедифференцировке SPA – VSMC.
Таким образом, мы изолировали первичные dNK и децидуальные макрофаги из тканей децидуальной оболочки человека на ранних сроках беременности с помощью магнитных шариков, специфичных для CD56 или CD14. Проточная цитометрия показала, что их чистота была более 90% (дополнительные рисунки S1B и C).Среды для кондиционирования dNK или децидуальных макрофагов собирали отдельно через 24 часа культивирования (dNK-CM и dM-CM) и использовали при 50% концентрации для обработки клеток T / G HA-VSMC в течение 3 дней. Как и ожидалось, уровни маркеров VSMC, αSMA, кальпонина, SM22α и MYh21 в клетках T / G HA-VSMC заметно снижались при обработке dNK-CM или dM-CM (Фигуры 4A и B и 5A и B). . Соответственно, изменение морфологии клеток до округлой формы и значительное уменьшение окрашивания αSMA в T / G HA-VSMC клетках, обработанных dNK-CM или dM-CM, были выявлены с помощью иммунофлуоресценции для αSMA (Фигуры 4C и 5C).Данные доказали активную функцию dNKs и децидуальных макрофагов, способствующих дедифференцировке VSMC.
Как указано выше, результаты анализа CCK8 (дополнительный рисунок S1E), проточной цитометрии для аннексина / PI и 7-AAD (дополнительный рисунок S2A и B) и вестерн-блоттинга для расщепленной каспазы 3 (дополнительный рисунок S2C) продемонстрировали, что лечение dNK -CM или dM-CM в T / G HA-VSMC не влияли на жизнеспособность клеток.
EVT улучшенная дедифференциация VSMC
Близкая локализация EVTs с SPA – VSMCs на стадии ремоделирования была хорошо известна, и здесь наши результаты иммунофлуоресценции при ремоделировании SPA подтвердили проявление (Figure 2D).В литературных исследованиях, посвященных влиянию клеток трофобласта на дедифференцировку SPA – VSMC, в качестве модели клеток обычно использовались ворсинчатые трофобласты [29, 30]. В то время как особенности ворсинчатых трофобластов сильно отличаются от децидуальных EVT. После этого мы отсортировали EVT в децидуальной оболочке на ранних сроках беременности с помощью магнитных шариков на HLA-G, который является специфическим маркером EVT. Проточная цитометрия показала, что чистота клеток составляла около 94% (дополнительный рисунок S1D). Клетки T / G HA-VSMC обрабатывали 50% кондиционной средой EVT (EVT-CM) в течение 3 дней.Неудивительно, что экспрессия αSMA, кальпонина, SM22α и MYh21 была явно подавлена обработкой (фиг. 6A и B), а фенотипы дедифференцировки были выявлены при иммуноокрашивании на αSMA (фиг. 6C).
Рисунок 6
Децидуальные EVT способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G, обработанных HA-VSMC с EVT-CTRL и EVT-CM ( n = 3 ).(C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с EVT-CTRL и EVT-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 6
Децидуальные EVT способствуют дедифференцировке VSMC. (A, B) Относительные уровни мРНК (A) и уровни белка (B) αSMA, кальпонина, SM22α или MYh21 в T / G, обработанных HA-VSMC с EVT-CTRL и EVT-CM ( n = 3 ).(C) Клеточное иммунофлуоресцентное окрашивание αSMA для совместных культур T / G HA-VSMC с EVT-CTRL и EVT-CM. Белыми стрелками обозначены клетки круглой формы. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Опять же, результаты анализа CCK8 (дополнительный рисунок S1E), проточной цитометрии для аннексина / PI и 7-AAD (дополнительный рисунок S2A и B) и вестерн-блоттинга для расщепленной каспазы 3 (дополнительный рисунок S2C) продемонстрировали, что лечение EVT- CM не влиял на жизнеспособность клеток в T / G HA-VSMC.
SPA – VSMC потенциально могут дедифференцироваться по отношению к dNK
Теперь, когда наши результаты подтвердили, что различные типы клеток в децидуальной оболочке участвуют в регуляции дедифференцировки SPA-VSMC, остается вопрос, где эти дедифференцированные клетки находятся. Примечательно, что тщательное исследование иммунофлуоресцентного окрашивания выявило наличие двойных положительных клеток SM22α и CD56, составляющих примерно 2% VSMC в области, близкой к ремоделирующимся SPA (рис. 7A), что указывает на то, что эти клетки могут обладать характеристиками как VSMC, так и dNKs.
Рисунок 7
SPA – VSMC потенциально дедифференцируются по отношению к dNK во время ремоделирования SPA. (A) Иммунофлуоресцентное окрашивание на SM22α (зеленый), CD56 (белый) и CK7 (красный) в децидуальной оболочке на ранних сроках беременности показывает наличие двойных положительных клеток SM22α и CD56 (стрелка). (B) Иммунофлуоресцентный анализ ДНК – ISH – PLA в децидуальной оболочке человека на ранних сроках беременности. Левая верхняя панель показывает типичный сигнал PLA (красный, стрелка) в VSMC, помеченных αSMA (зеленый). Правая верхняя панель показывает сигнал PLA (красный, стрелка) в отдельных dNK, который отмечен CD56 (зеленый).Левая нижняя панель показывает несколько сигналов PLA (красный) в EVT, которые помечены CK7 (зеленый). Правая нижняя панель показывает несколько сигналов PLA (красный) в DSC, которые помечены Vimentin (зеленый). (C) Статистические результаты анализа ChIP в CD56 + dNK, показывающие обогащение h4K4dime в промоторной области гена MYh21 . Статистические результаты анализа ChIP в EVT и DSC в качестве отрицательного контроля показывают меньшее обогащение h4K4dime в промоторной области гена MYh21 . Статистический анализ проводится по не менее чем трем независимо повторяющимся экспериментам.Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Рисунок 7
SPA – VSMC потенциально дедифференцируются в сторону dNK во время ремоделирования SPA. (A) Иммунофлуоресцентное окрашивание на SM22α (зеленый), CD56 (белый) и CK7 (красный) в децидуальной оболочке на ранних сроках беременности показывает наличие двойных положительных клеток SM22α и CD56 (стрелка). (B) Иммунофлуоресцентный анализ ДНК – ISH – PLA в децидуальной оболочке человека на ранних сроках беременности.Левая верхняя панель показывает типичный сигнал PLA (красный, стрелка) в VSMC, помеченных αSMA (зеленый). Правая верхняя панель показывает сигнал PLA (красный, стрелка) в отдельных dNK, который отмечен CD56 (зеленый). Левая нижняя панель показывает несколько сигналов PLA (красный) в EVT, которые помечены CK7 (зеленый). Правая нижняя панель показывает несколько сигналов PLA (красный) в DSC, которые помечены Vimentin (зеленый). (C) Статистические результаты анализа ChIP в CD56 + dNK, показывающие обогащение h4K4dime в промоторной области гена MYh21 .Статистические результаты анализа ChIP в EVT и DSC в качестве отрицательного контроля показывают меньшее обогащение h4K4dime в промоторной области гена MYh21 . Статистический анализ проводится по не менее чем трем независимо повторяющимся экспериментам. Данные представлены в виде среднего значения ± стандартное отклонение, и сравнение между группами выполняется с помощью критерия Стьюдента t . * P <0,05. Шкала показывает 100 мкм.
Сообщалось, что VSMC специфически проявляют модификацию h4K4dime на промоторе гена MYh21 [19, 31], что может быть обнаружено с помощью анализа IF-ISH-PLA [19].Этот метод идентифицирует h4K4dime в промоторе MYh21 по сигналу PLA в отдельной клетке, которая помечена иммунофлуоресценцией для определенного клеточного маркера. Используя этот метод, мы обнаружили в децидуальной ткани человека примерно 5% клеток CD56 + dNK отображали положительный сигнал для PLA, который обычно обнаруживался в αSMA-положительных SPA – VSMC (рис. 7B). Однако ни EVT, меченные СК, ни ДСК, меченные виментином, не показали никакого сигнала PLA (рис. 7B). Данные показали, что небольшое количество клеток CD56 + dNK специфически проявляет модификацию h4K4dime в промоторной области MYh21 .
Для подтверждения результатов первичные dNK, EVT и DSC, очищенные из децидуальной оболочки человека на ранних сроках беременности (дополнительный рисунок S1), подвергали анализу ChIP. Результаты выявили очевидную модификацию h4K4dime в промоторной области MYh21 в dNK, но не в EVT или DSC (фиг. 7C). Взятые вместе, эти результаты подтверждают, что часть SPA-VSMCs может дедифференцироваться в сторону dNKs во время ремоделирования SPA.
Обсуждение
Успех беременности регулируется сложным взаимодействием клеток на границе раздела матери и плода, и многие клетки динамически изменяли свои характеристики и функции, чтобы удовлетворить потребность в росте плода.Физиологические изменения SPA – VSMCs во время ремоделирования SPA отражают сложные перекрестные помехи между множественными типами клеток. Обобщая данные этого исследования и литературные данные, можно сказать, что процесс дедифференцировки SPA-VSMCs проиллюстрирован на рисунке 8. В небеременной матке очень мало лимфоцитов существует в эндометрии [32, 33], а SPA эндометрия имеют интактный слой VSMC в какие SPA – VSMC представляют типичную функцию SMC [4, 20] (рисунок 8A). С физиологической точки зрения децидуализация эндометрия матки является преобладающим процессом подготовки эндометрия к беременности.У человека процесс децидуализации запускается в середине секреторной фазы менструального цикла и широко проявляется во время беременности [34, 35]. Важно отметить, что децидуализация происходит сначала вокруг терминальных СПА поверхностного слоя эндометрия, и в конечном итоге она затрагивает весь эндометрий во время беременности. Хорошо известно, что активация пути цАМФ, который может повышать чувствительность эндометриальных клеток человека к прогестерону, является начальным и обязательным событием, запускающим децидуальный процесс [36].После эмбриональной имплантации огромное количество полученного из трофобласта человеческого хорионического гонадотропина (ХГЧ) передает сигнал через путь цАМФ, преимущественно ускоряя процесс децидуализации [37]. DSC активны в производстве большого количества цитокинов и хемокинов, которые имеют решающее значение для регистрации и модуляции лимфоцитов и, таким образом, для защиты генетически «чужеродного» полуаллогенного эмбриона. Морфологические изменения SPA-VSMC в артериях Un-Rem и заметные изменения в клеточных свойствах VSMC при обработке условной средой из DSC, dNK или децидуальных макрофагов указывают на первичную роль DSC в инициации дедифференцировки SPA-VSMC, которая впоследствии усиливается этими лимфоцитами перед участием EVT (рис. 8B и C).Драматическая дедифференцировка SPA-VSMCs происходит после вторжения EVTs, в конечном итоге приводя к исчезновению VSMCs на стадии Fully-Rem (Рис. 8D и E). При доношенной беременности, когда полные SPA были реконструированы, EVT в диком виде существовали вокруг и внутри SPA, а SPA – VSMC почти полностью исчезли (дополнительный рисунок S3). Каскадная регуляция дедифференцировки SPA-VSMC отражает гармонизированные взаимодействия между множественными типами клеток в децидуальном компартменте.
Рисунок 8
Схематический рисунок, иллюстрирующий постепенное преобразование SPA – VSMC во время ремоделирования SPA.(A) На небеременной стадии SPA – VSMC эндометрия остаются неизменными в менструальном цикле, и небольшое количество dNK и децидуальных макрофагов существует в эндометрии. (B) Стромальные клетки эндометрия подвергаются децидуализации при имплантации эмбриона, и они инициируют морфологическую трансформацию SPA – VSMCs. (C) Децидуальные NKs и децидуальные макрофаги рекрутируются в децидуальный компартмент, и они усиливают разделение, смещение и округление SPA – VSMCs на стадии Un-Rem. (D) Экстраворсинчатые трофобласты вторгаются и работают вместе с dNKs и децидуальными макрофагами, чтобы вызвать заметную дедифференцировку SPA-VSMCs на стадии ремоделирования.Обратите внимание на появление клеток с двойной функцией VSMC и dNKs, окружающих ремоделирующуюся SPA (розовые клетки). (E) VSMC спиральной артерии исчезают на стадии Fully-Rem.
Рисунок 8
Схематический рисунок, иллюстрирующий постепенное преобразование SPA – VSMC во время ремоделирования SPA. (A) На небеременной стадии SPA – VSMC эндометрия остаются неизменными в менструальном цикле, и небольшое количество dNK и децидуальных макрофагов существует в эндометрии. (B) Стромальные клетки эндометрия подвергаются децидуализации при имплантации эмбриона, и они инициируют морфологическую трансформацию SPA – VSMCs.(C) Децидуальные NKs и децидуальные макрофаги рекрутируются в децидуальный компартмент, и они усиливают разделение, смещение и округление SPA – VSMCs на стадии Un-Rem. (D) Экстраворсинчатые трофобласты вторгаются и работают вместе с dNKs и децидуальными макрофагами, чтобы вызвать заметную дедифференцировку SPA-VSMCs на стадии ремоделирования. Обратите внимание на появление клеток с двойной функцией VSMC и dNKs, окружающих ремоделирующуюся SPA (розовые клетки). (E) VSMC спиральной артерии исчезают на стадии Fully-Rem.
Ремоделирование SPA перед инвазией клеток трофобласта является предпосылкой для следующего ремоделирования EVT маточных артерий. Раннее исследование гистерэктомии показало, что клетки трофобласта не могут проникать в стенку маточного SPA на небеременной стадии, но они могут это сделать в образцах матки, взятых с помощью кесарева сечения при доношенной беременности [38]. Безусловно, многие исследования выяснили действие лимфоцитов матки, в первую очередь dNKs и макрофагов, в индукции трансформации или дедифференцировки VSMC матки [8, 39, 40].В этом пункте мы сообщили о результатах, аналогичных предыдущим исследованиям. Однако остается неизвестным, участвуют ли и каким образом децидуальные клетки стромы в этом процессе. Недавний одноклеточный анализ взаимодействия матери и плода человека на ранних сроках беременности аннотировал молекулярные особенности трех подмножеств ДСК, а анализ взаимодействия лиганд-рецептор позволил предположить потенциальное перекрестное взаимодействие подмножеств SC в компактной децидуальной оболочке с периваскулярными SMC через IGF-IGF1R , Распознавание VEGF-KDR / NRP1 [41]. Кроме того, исследования артерий из других тканей показали, что PDGF-BB играет критическую роль в обеспечении дедифференцировки VSMC посредством передачи сигналов Erk [42].О продукции PDGF с помощью DSC сообщалось в нескольких исследованиях [43–45]. Эти доказательства, вместе с тем фактом, что маточные артерии анатомически встроены в строму, указывают на способность DSCs вызывать начальный сигнал, индуцирующий дедифференцировку SPA-VSMCs. Действительно, наши данные in vitro о клетках VSMC предоставляют убедительные доказательства такой функции DSC, хотя рабочие факторы еще предстоит идентифицировать.
События децидуализации стромы, инфильтрации лимфоцитов, инвазии EVT и замещения эндотелия происходят по очереди во время беременности.Анализ одноклеточного секвенирования децидуальной оболочки человека обеспечивает надежные молекулярные доказательства сложного взаимодействия между DSC, иммунными клетками, EVT, эндотелиальными клетками и периваскулярными SMC [41]. Накапливающиеся доказательства показывают, что большое количество иммунных клеток рекрутируется одновременно с децидуализацией [46], и множественные децидуальные хемокины, такие как CXCL6 и CCL14, имеют решающее значение для накопления dNKs и макрофагов в области, окружающей SPA [13]. Привлеченные иммунные клетки работают вместе с децидуальными клетками, направляя инвазию трофобластов и ремоделируя артерии [26, 47].С другой стороны, инвазивные EVT играют ключевую роль в снижении иммунно-атакующих свойств лимфоцитов [48]. Важно отметить, что активная интермодуляция между различными иммунными клетками гарантирует смещенную иммунно-защитную среду в децидуальной оболочке. В сложном децидуальном компартменте MMP9, ангиопоэтин (Ang) -1, Ang-2, интерферон-g, VEGF-C из клеток dNK [40, 9], IP-10 и CXCL10 из EVT [49] и фагоцитотическая функция Макрофаги [40], как было показано, в значительной степени нарушают целостность VSMC и ингибируют экспрессию маркерного гена VSMC [49].Помимо этих факторов, плацента активно продуцирует многочисленные miRNAs, которые, как было доказано, участвуют в модуляции различных клеточных событий, включая инвазию клеток трофобласта и ремоделирование SPA [50, 51]. Напр., Происходящий из плаценты miR-210 может нацеливаться на Efna3 и Bcl2, которые участвуют в ангиогенезе и трансформации VSMCs [52–55]. Кроме того, ряд исследований демонстрирует, что VSMC и эндотелий SPA экспрессируют хемокины, включая CCL4, 14, 16, 21, 22, CXCL12 и CX3CR1 [7, 13, 56-59], которые могут специфически привлекать макрофаги и NK и регулируют вторжение трофобластов.Следовательно, сложная регуляторная сеть среди DSC, децидуальных лейкоцитов, EVT, VSMC и эндотелия может определять успех ремоделирования сосудов. Сложность микросреды также отражена в наблюдении, что децидуальные СПА демонстрируют несинхронизированные фенотипические изменения на ранних сроках беременности. Состав типов клеток вокруг отдельных SPA варьируется, и дедифференцировка VSMC кажется более интенсивной в SPA, окруженной как EVT, так и иммунными клетками, по сравнению с этим SPA без EVT.Следовательно, стоит продолжить изучение того, как множественные клетки взаимодействуют пространственно-временным динамическим и гармонизированным образом, чтобы регулировать дедифференцировку SPA-VSMC.
На сегодняшний день судьба дедифференцированных SPA – VSMC остается загадкой. Наше исследование предоставляет первые доказательства потенциальной трансформации SPA – VSMC в CD56 + dNK. Модификация h4K4dime в промоторной области MYh21 в небольшой части dNK указывает на их происхождение от VSMC. Те клетки, окружающие ремоделирующие артерии с коэкспрессией SM22 и CD56, могут быть SPA-VSMCs, которые дедифференцируются в сторону dNK.Это открытие представляет особый интерес для изучения особого происхождения клеток dNK. Делеция T-bet и Nfil3, двух важных факторов транскрипции, управляющих развитием NK-клеток у мышей, серьезно снижает функциональные NK-клетки в других органах, но мало влияет на обогащение NK матки во время беременности [60–62]. Поэтому считается, что NK-клетки матки могут иметь различное происхождение. Здесь наши данные предполагают, что небольшая часть dNK потенциально происходит от VSMC.Пластичность SMCs была признана, и они могут дедифференцироваться до макрофагов, остеохондроцитов или адипоцитов при различных патологических состояниях [63–65]. Доказательства из модели мыши с отслеживанием клонов окончательно показали критическую роль KLF4 в регуляции фенотипических переходов SMCs [65]. Интересно, что исследования в матке демонстрируют участие KLF4 в регуляции рецептивности матки, что проявляется в стимуляции репликации эпителиальной ДНК с помощью эстроген-индуцированного KLF4, в то время как существует ингибирование этого процесса с помощью индуцированного прогестероном KLF15 [66].Хорошо известно, что маточные СПА чувствительны к стероидам [67–71]. Следовательно, наиболее вероятно, что гормоны беременных, эстроген и прогестерон, могут индуцировать факторы транскрипции, включая KLF4 и KLF15, в SPA-VSMC и регулировать трансформацию этих клеток в dNKs или другие типы клеток. С другой стороны, трансформация SPA-VSMCs в dNK также указывает на прямой механизм дедифференцировки VSMC во время ремоделирования сосудов матки. Основные механизмы требуют дальнейшего изучения.
Хотя мы наблюдали значительное снижение экспрессии маркерного гена VSMC и морфологические изменения в T / G HA-VSMC, которые обрабатывались кондиционирующей средой из различных децидуальных клеток, мы не обнаружили индукции маркеров dNK (дополнительный рисунок S4). Наиболее вероятно, что клетки T / G HA-VSMC не обладают реальными характеристиками SPA-VSMC. Свойства VSMC из разных органов или из разных сред различны [72, 73]. Модели in vitro, такие как совместное культивирование эксплантата плаценты и децидуальной оболочки, совместное культивирование EVTs-эндотелиальных клеток и VSMCs или культура хорионической пластинчатой артерии (CPA), были использованы для изучения механизмов ремоделирования сосудов матки человека.Несомненно, что первичные SPA – VSMC из эндометрия человека на небеременной стадии или на ранних сроках беременности будут оптимальной моделью in vitro. Однако получить образцы SPA относительно сложно из-за их довольно небольшого размера (дополнительный рисунок S5). Улучшение методологии будет необходимо для получения достаточного количества SPA – VSMC для дальнейшего изучения механизма. Другой проблемой является создание оптимизированных условий индукции, имитирующих микросреду вокруг SPA – VSMC. Подходящая комбинация кондиционированных сред или факторов из разных типов децидуальных клеток может работать более эффективно, чтобы вызвать дедифференцировку VSMC.Однако это критически зависит от всестороннего определения факторов из различных типов клеток в нише децидуальной SPA в разных точках беременности.
В целом, наши данные указывают на сигнальный каскад от DSCs, dNKs, децидуальных макрофагов и EVTs, чтобы вызвать прогрессивную дедифференцировку маточных SPA – VSMCs и выявить потенциальную трансформацию SPA – VSMC в dNKs. Полученные данные расширяют наше понимание неотъемлемых ролей децидуальной ниши, которые контролируют дедифференцировку SPA – VSMC.Углубленное исследование основных механизмов может помочь в разработке новых стратегий предотвращения неблагоприятных исходов беременности, связанных с неудачами ремоделирования сосудов, такими как преэклампсия и выкидыш.
Благодарность
Благодарим за техническую поддержку г-жи Шивен Ли, Сили Чжу и Ся Ян в экспериментах по конфокальному анализу и анализу проточной цитометрии.
Конфликт интересов
Авторы заявили об отсутствии конфликта интересов.
Вклад авторов
Ю.М. провели эксперименты, интерпретировали данные и составили рукопись. X.Y. и Л.З. собрали клинические образцы, подготовили разделы и интерпретировали данные. J.L. X.S. и Y.-x.L. помог в сборе образцов и культивировании клеток. Y.L.W. разработал и контролировал исследование, интерпретировал данные и редактировал рукопись.
Список литературы
1.Королевство
JC
,Kaufmann
P
.Кислород и развитие сосудов плаценты
.Adv Exp Med Biol
1999
;474
:259
—275
.2.Плеть
GE
,Эрнеруд
J
.Децидуальные цитокины и осложнения беременности: внимание к самопроизвольному выкидышу
.J Репрод Иммунол
2015
;108
:83
—89
.3.Burton
GJ
,Jauniaux
E
.Патофизиология ограничения роста плода из-за плаценты
.Am J Obstet Gynecol
2018
;218
:S745
—S761
. 4.Pijnenborg
R
,Vercruysse
L
,Hanssens
M
.Спиральные артерии матки при беременности человека: факты и противоречия
.Плацента
2006
;27
:939
—958
.5.Harris
LK
,Keogh
RJ
,Wareing
M
,Baker
PN
,Cartwright
JE
,000
GS5000 Whitley
.Инвазивные трофобласты стимулируют апоптоз гладкомышечных клеток сосудов с помощью fas-лиганд-зависимого механизма
.Am J Pathol
2006
;169
:1863
—1874
.6.Keogh
RJ
,Harris
LK
,Freeman
A
,Baker
PN
,Aplin
JD
, Whitley.Трофобласты плода используют лиганд, индуцирующий апоптоз, связанный с апоптотическим цитокином фактора некроза опухоли альфа, для индукции гибели гладкомышечных клеток
.Circ Res
2007
;100
:834
—841
.7.Smith
SD
,Dunk
CE
,Aplin
JD
,Harris
LK
,Jones
RL
.Доказательства участия иммунных клеток в ремоделировании децидуальной спиральной артериолы на ранних сроках беременности человека
.Am J Pathol
2009
;174
:1959
—1971
.8.Bulmer
JN
,Innes
BA
,Levey
J
,Robson
SC
,Lash
GE
.Роль апоптоза и миграции клеток гладких мышц сосудов во время ремоделирования маточной спиральной артерии при нормальной беременности человека
.FASEB J
2012
;26
:2975
—2985
.9.Robson
A
,Harris
LK
,Innes
BA
,Lash
GE
,Aljunaidy
MM
,0000004 AplinAplin
Робсон
SC
,Bulmer
JN
.Естественные клетки-киллеры матки инициируют ремоделирование спиральной артерии при беременности человека
.FASEB J
2012
;26
:4876
—4885
.10.Gomez
D
,Owens
GK
.Фенотипическое переключение гладкомышечных клеток при атеросклерозе
.Cardiovasc Res
2012
;95
:156
—164
. 11.Misra
A
,Sheikh
AQ
,Kumar
A
,Luo
J
,Zhang
J
,000 RB 9000 9000 9000 Smoot4 9000 9000 Smoot4 9000Каплан
P
,Городской
Z
,Qyang
Y
,Tellides
G
,Greif
DM
.Ингибирование интегрина бета3 — терапевтическая стратегия надклапанного стеноза аорты
.J Exp Med
2016
;213
:451
—463
.12.Zhu
H
,Hou
CC
,Luo
LF
,Hu
YJ
,Yang
WX
.Стромальные клетки эндометрия и децидуализированные стромальные клетки: происхождение, трансформация и функции
.Gene
2014
;551
:1
—14
. 13.Choudhury
RH
,Dunk
CE
,Lye
SJ
,Aplin
JD
,Harris
LK
,Jones
Взаимное взаимодействие вневорсинчатых трофобластов и эндотелиальных клеток опосредует инфильтрацию лейкоцитов для раннего ремоделирования стенки децидуальной спиральной артериолы
.J Immunol
2017
;198
:4115
—4128
. 14.Faas
MM
,de
Vos
P
.NK-клетки и макрофаги матки при беременности
.Плацента
2017
;56
:44
—52
. 15.Wu
HM
,Huang
HY
,Soong
YK
,Leung
PCK
,Wang
HS
.Кисспептин регуляция подвижности децидуальных стромальных клеток человека посредством внутриклеточных тирозинкиназ FAK-Src
.Hum Reprod
2019
;34
:1291
—1301
. 16.млн. Лет
L
,Li
G
,Cao
G
,Zhu
Y
,Du
MR
,Zhao
Y
Лю
Y
,Ян
Y
,Li
YX
,Li
DJ
,Ян
H
и др. КлеткиdNK способствуют взаимодействию между трофобластическими и эндотелиальными клетками через VEGF-C и HGF
.Immunol Cell Biol
2017
;95
:695
—704
. 17.Sayama
S
,Nagamatsu
T
,Schust
DJ
,Itaoka
N
,Ichikawa
M
,000000 Kaw4000,000 Kaw4000 Kaw4000Kozuma
S
,Fujii
T
.Децидуальные макрофаги человека подавляют продукцию IFN-гамма Т-клетками посредством костимулирующей передачи сигналов B7-h2: PD-1 на ранних сроках беременности
.J Репрод Иммунол
2013
;100
:109
—117
. 18.Hanna
J
,Goldman-Wohl
D
,Hamani
Y
,Avraham
I
,Greenfield
C
,Natanson-Yar
D
,Cohen-Daniel
L
,Arnon
TI
,Manaster
I
,Gazit
R
,Yutkin
V
и др.Децидуальные NK-клетки регулируют ключевые процессы развития на стыке плода и матери человека
.Nat Med
2006
;12
:1065
—1074
.19.Gomez
D
,Shankman
LS
,Nguyen
AT
,Owens
GK
.Обнаружение модификаций гистонов в определенных локусах генов в отдельных клетках в гистологических срезах
.Nat Methods
2013
;10
:171
—177
.20.Biswas Shivhare
S
,Bulmer
JN
,Innes
BA
,Hapangama
DK
,Lash
GE
.Измененная дифференцировка гладкомышечных клеток сосудов в сосудистой сети эндометрия при меноррагиях
.Hum Reprod
2014
;29
:1884
—1894
.21.Ramathal
CY
,Bagchi
IC
,Taylor
RN
,Bagchi
MK
.Децидуализация эндометрия: мышей и мужчин
.Semin Reprod Med
2010
;28
:17
—26
. 22.Ashary
N
,Tiwari
A
,Modi
D
.Имплантация эмбриона: война во времена любви
.Эндокринология
2018
;159
:1188
—1198
. 23.Наружный
V
,Trundley
A
,Gardner
L
,Northfield
J
,Chang
C
,Apps
R
R
Естественные клетки-киллеры при беременности человека
.Методы Мол Биол
2010
;612
:447
—463
.24.Моффет-Кинг
А
.Естественные клетки-киллеры и беременность
.Nat Rev Immunol
2002
;2
:656
—663
0,25.Vacca
P
,Mingari
MC
,Moretta
L
.Естественные клетки-киллеры при беременности человека
.J Репрод Иммунол
2013
;97
:14
—19
.26.Liu
S
,Diao
L
,Huang
C
,Li
Y
,Zeng
Y
,H
J Kwak-KimРоль децидуальных иммунных клеток в беременности человека
.J Репрод Иммунол
2017
;124
:44
—53
. 27.Сойка
ДК
,Ян
L
,Йокояма
WM
.Естественные киллеры матки
.Фронт Иммунол
2019
;10
:960
. 28.Манастер
I
,Мандельбойм
O
.Уникальные свойства человеческих NK-клеток слизистой оболочки матки
.Плацента
2008
;29
:S60
—S66
. 29.Red-Horse
K
,Zhou
Y
,Genbacev
O
,Prakobphol
A
,Foulk
R
,McMaster
McMaster
.Дифференциация трофобластов во время имплантации эмбриона и формирование границы раздела матери и плода
.J Clin Invest
2004
;114
:744
—754
. 30.Харрис
LK
,Джонс
CJ
,Аплин
JD
.Молекулы адгезии в трофобласте человека — обзор. II. Трофобласт вневорсинчатый
.Плацента
2009
;30
:299
—304
.31.McDonald
OG
,Wamhoff
BR
,Hoofnagle
MH
,Owens
GK
.Контроль связывания SRF с хроматином CArG-бокса регулирует экспрессию генов гладких мышц in vivo
.J Clin Invest
2006
;116
:36
—48
. 32.Брайтон
PJ
,Маруяма
Y
,Fishwick
K
,Vrljicak
P
,Tewary
S
,0004 Fuji
Lucas
ES
,Yamada
T
,Woods
L
,Lucciola
R
,Hou Lee
Y
et al.Удаление стареющих децидуальных клеток маточными естественными клетками-киллерами в циклическом эндометрии человека
.Элиф
2017
;6
: e31274.33.Thiruchelvam
U
,Dransfield
I
,Saunders
PT
,Critchley
HO
.Важность макрофагов в эндометрии человека
.J Leukoc Biol
2013
;93
:217
—225
.34.Edgell
TA
,Rombauts
LJ
,Salamonsen
LA
.Оценка восприимчивости эндометрия: необходимость быстрого неинвазивного теста
.Reprod Biomed Online
2013
;27
:486
—496
0,35.Salamonsen
LA
,Evans
J
,Nguyen
HP
,Edgell
TA
.Микросреда имплантации человека: фактор репродуктивного успеха
.Am J Reprod Immunol
2016
;75
:218
—225
.36.Brar
AK
,Frank
GR
,Kessler
CA
,Cedars
MI
,Handwerger
S
.Прогестерон-зависимая децидуализация эндометрия человека опосредуется цАМФ
.Эндокринная
1997
;6
:301
—307
0,37.Weedon-Fekjaer
MS
,Tasken
K
.Обзор: пространственно-временная динамика передачи сигналов ХГЧ / цАМФ и регуляция функции плаценты
.Плацента
2012
;33
:S87
—S91
. 38.Crocker
IP
,Wareing
M
,Ferris
GR
,Jones
CJ
,Cartwright
JE
,JE
,0005Влияние сосудистого происхождения, кислорода и фактора некроза опухоли альфа на трофобластную инвазию материнских артерий in vitro
.J Pathol
2005
;206
:476
—485
. 39.Robson
A
,Lash
GE
,Innes
BA
,Zhang
JY
,Robson
SC
,0004 BulmerДедифференцировка мышц спиральной артерии матки
.Hum Reprod
2019
;34
:1428
—1438
.40.Hazan
AD
,Smith
SD
,Jones
RL
,Whittle
W
,Lye
SJ
,Dunk CE
.
Сосудисто-лейкоцитарные взаимодействия: механизмы ремоделирования децидуальной спиральной артерии человека in vitro
.Am J Pathol
2010
;177
:1017
—1030
.41.Vento-Tormo
R
,Efremova
M
,Botting
RA
,Turco
MY
,Vento-Tormo
M
KB Park
JE
,Stephenson
E
,Polanski
K
,Goncalves
A
,Gardner
L
,Holmqvist
et al.
Одноклеточная реконструкция ранней границы раздела матери и плода у человека
.Природа
2018
;563
:347
—353
.42.Dong
LH
,Wen
JK
,Miao
SB
,Jia
Z
,Hu
HJ
,RR
, Sun
Хан
М
.Байкалин ингибирует стимулированную PDGF-BB пролиферацию гладкомышечных клеток сосудов посредством подавления передачи сигналов PDGFRbeta-ERK и увеличения накопления p27 и предотвращает индуцированную повреждением гиперплазию неоинтимы
.Cell Res
2010
;20
:1252
—1262
. 43.Kingsley
K
,Huff
JL
,Rust
WL
,Carroll
K
,Martinez
AM
,000 Plopper
000 Plopper000ERK1 / 2 опосредует PDGF-BB стимулированную пролиферацию и миграцию клеток гладких мышц сосудов на ламинине-5
.Biochem Biophys Res Commun
2002
;293
:1000
—1006
. 44.юаней
X
,Zhang
T
,Yao
F
,Liao
Y
,Liu
F
,Ren
ZhangZ
Diao
L
,Li
Y
,Zhou
B
,He
F
,Wang
L
.THO комплекс-зависимый посттранскрипционный контроль вносит вклад в решение судьбы гладкомышечных клеток сосудов
.Circ Res
2018
;123
:538
—549
. 45.Saji
M
,Taga
M
,Matsui
H
,Suyama
K
,Kurogi
K
,Minaguchi
9.Экспрессия генов и специфическое связывание тромбоцитарного фактора роста и его влияние на синтез ДНК в децидуальных клетках человека
.Mol Cell Endocrinol
1997
;132
:73
—80
. 46.Lockwood
CJ
,Huang
SJ
,Chen
CP
,Huang
Y
,Xu
J
,Kayarz 9000
000000 SJKayisli
U
,Koopman
L
,Smedts
D
,Buchwalder
LF
,Schatz
F
.Децидуальная клеточная регуляция естественных хемокинов, рекрутирующих клетки-киллеры: значение для патогенеза и прогнозирования преэклампсии
.Am J Pathol
2013
;183
:841
—856
. 47.Wallace
AE
,Fraser
R
,Cartwright
JE
.Экстраворсинчатый трофобласт и децидуальные естественные клетки-киллеры: ремоделирующее партнерство
.Hum Reprod Update
2012
;18
:458
—471
. 48.Wu
X
,Jin
LP
,юаней
MM
,Zhu
Y
,Wang
MY
,Li
DJ
DJ
.Человеческие клетки трофобласта первого триместра привлекают CD56brightCD16- NK-клетки в децидуальную оболочку посредством экспрессии и секреции фактора 1 CXCL12 / стромальных клеток
.J Immunol
2005
;175
:61
—68
. 49.Wallace
AE
,Cartwright
JE
,Begum
R
,Laing
K
,Thilaganathan
B
, WhitИндуцированные трофобластом изменения в экспрессии хемокина 10 мотива C-x-C способствуют дедифференцировке гладкомышечных клеток сосудов во время ремоделирования спиральной артерии
.Артериосклер Тромб Васк Биол
2013
;33
:e93
—e101
.50.Askie
LM
,Duley
L
,Henderson-Smart
DJ
,Stewart
LA
,Group PC
.Антиагреганты для профилактики преэклампсии: метаанализ индивидуальных данных пациента
.Ланцет
2007
;369
:1791
—1798
.51.Ляо
Y
,Lonnerdal
B
.miR-584 опосредует посттранскрипционную экспрессию рецептора лактоферрина в клетках Caco-2 и в тонком кишечнике мыши в перинатальный период
.Int J Biochem Cell Biol
2010
;42
:1363
—1369
. 52.Sun
Y
,X
,Liu
Q
,Wang
Z
,Xin
Y
,Zhang
000,000 P4Лю
Y
.Аутофагия, индуцированная гипоксией, снижает радиочувствительность пути HIF-1alpha / miR-210 / Bcl-2 в раковых клетках толстой кишки
.Int J Oncol
2015
;46
:750
—756
. 53.Bao
Q
,Jia
H
,Rong
A
,Cao
Z
,Zhang
Y
.MiR-210 ингибирует индуцированный гипоксией апоптоз гладкомышечных клеток посредством нацеливания на MEF2C
.Int J Clin Exp Pathol
2019
;12
:1846
—1858
. 54.Кхонг
TY
,Абдул Рахман
H
.Экспрессия Bcl-2 задерживает послеродовую инволюцию вызванных беременностью сосудистых изменений в плацентарном ложе человека
.Int J Gynecol Pathol
1997
;16
:138
—142
.55.Wang
H
,Wang
L
,Zhou
X
,Luo
X
,Liu
K
,Jiang
E4Шао
Z
,Шан
Z
.Экзосомы OSCC регулируют miR-210-3p, направленную на EFNA3, чтобы способствовать ангиогенезу рака полости рта через путь PI3K / AKT
.Biomed Res Int
2020
;2020
: 2125656.56.Hannan
NJ
,Jones
RL
,Белый
CA
,Salamonsen
LA
.Хемокины, CX3CL1, CCL14 и CCL4, способствуют миграции трофобластов человека на границе раздела между плодом и матерью
.Biol Reprod
2006
;74
:896
—904
. 57.Jones
RL
,Hannan
NJ
,Kaitu’u
TJ
,Zhang
J
,Salamonsen
LA
.Идентификация хемокинов, важных для рекрутирования лейкоцитов в эндометрий человека во время имплантации эмбриона и менструации
.J Clin Endocrinol Metab
2004
;89
:6155
—6167
.58.Lu
H
,Jin
LP
,Huang
HL
,Ha
SY
,Yang
HL
,Chang
DJ5
,Li
MQ
.CXCL12, происходящий из трофобластов, способствует обогащению CD56 (ярких) CD82 (-) CD29 (+) NK-клеток в децидуальной оболочке
.Am J Reprod Immunol
2020
;83
: e13203.59.Smith
SDAJ
,Harris
LK
,Dunk
C
,Jones
RL
.Возможное участие хемокинов в рекрутировании клеток uNK и макрофагов для ремоделирования спиральных артерий
.Reprod Sci
2009
;16
:110A
.60.Tayade
C
,Клык
Y
,Черный
GP
,VA
PJ
,Erlebacher
A
,Croy
BA
Дифференциальная транскрипция Eomes и T-bet во время созревания естественных клеток-киллеров матки мыши
.J Leukoc Biol
2005
;78
:1347
—1355
.61.Рыжий
ML
,Portilho
NA
,Felker
AM
,Mohammad
S
,Mara
DL
,Croy
.Фактор транскрипции NFIL3 необходим для нормального плацентарного и эмбрионального развития, но не для дифференцировки клеток естественных киллеров матки (UNK) у мышей
.Biol Reprod
2016
;94
:101
.62.Sojka
DK
,Plougastel-Douglas
B
,Yang
L
,Pak-Wittel
MA
,Артемов
0004 MNova
C
,Chase
JM
,Rothman
PB
,Yu
J
,Riley
JK
,Zhu
J
et al.Резидентные в тканях естественные клетки-киллеры (NK) представляют собой линии клеток, отличные от NK-клеток тимуса и селезенки
.Элиф
2014
;3
:e01659
.63.длинный
JZ
,Svensson
KJ
,Tsai
L
,Zeng
X
,Roh
HC
,Kong
X4
Lou
J
,Lokurkar
I
,Baur
W
,Castellot
JJ
Jr,Rosen
ED
et al.Гладкомышечное происхождение бежевых адипоцитов
.Cell Metab
2014
;19
:810
—820
.64.Speer
MY
,Yang
HY
,Brabb
T
,Leaf
E
,Look
A
,Lin
WL4
WL4 WL4
Дичек
D
,Giachelli
CM
.Гладкомышечные клетки дают начало остеохондрогенным предшественникам и хондроцитам в кальцифицирующих артериях
.Circ Res
2009
;104
:733
—741
0,65.Shankman
LS
,Gomez
D
,Cherepanova
OA
,Salmon
M
,Alencar
GF
000000 Paskka000000 PaskkaNewman
AA
,Greene
ES
,Straub
AC
,Isakson
B
,Randolph
GJ
et al.KLF4-зависимая фенотипическая модуляция гладкомышечных клеток играет ключевую роль в патогенезе атеросклеротических бляшек
.Nat Med
2015
;21
:628
—637
0,66.Bhurke
AS
,Bagchi
IC
,Bagchi
MK
.Регулируемые прогестероном факторы эндометрия, контролирующие имплантацию
.Am J Reprod Immunol
2016
;75
:237
—245
.67.Maliqueo
M
,Echiburu
B
,Crisosto
N
.Половые стероиды модулируют маточно-плацентарную сосудистую систему: значение для акушерства и неонатальных исходов
.Front Physiol
2016
;7
:152
.68.Дики
RP
,Hower
JF
.Взаимосвязь уровней эстрадиола и прогестерона с кровотоком в матке на ранних сроках беременности
.Ранняя беременность
1996
;2
:113
—120
0,69.Альбрехт
ED
,Bonagura
TW
,Burleigh
DW
,Эндерс
AC
,Aberdeen
GW
000000 GW0000004 PepeПодавление эстрогеном инвазии вневорсинчатыми трофобластами спиральных артерий матки во время ранней беременности бабуина
.Плацента
2006
;27
:483
—490
.70.Bonagura
TW
,Babischkin
JS
,Aberdeen
GW
,Pepe
GJ
,Albrecht
ED
.Преждевременное повышение уровня эстрадиола на ранних сроках беременности павиана подавляет ремоделирование маточной артерии и экспрессию экстравирусного фактора роста эндотелия сосудов плаценты и интегринов альфа1бета1 и альфа5бета1
.Эндокринология
2012
;153
:2897
—2906
. 71.Aberdeen
GW
,Bonagura
TW
,Harman
CR
,Pepe
GJ
,Albrecht
ED
.Подавление эстрогеном ремоделирования спиральной артерии матки трофобласта при беременности павианом: влияние на динамику кровотока матки и плода
.Am J Physiol Heart Circ Physiol
2012
;302
:h2936
—h2944
.72.млн.
.Сравнение электрофизиологических свойств гладкомышечных клеток сосудов в различных артериолах морских свинок
.Шэн Ли Сюэ Бао
2010
;62
:421
—426
.73.Hill
MA
,Meininger
GA
.Клетки гладких мышц артериолярных сосудов: механотрансдукторы в сложной среде
.Int J Biochem Cell Biol
2012
;44
:1505
—1510
.Заметки автора
© Автор (ы) 2020. Опубликовано Oxford University Press от имени Общества изучения репродукции.Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
Анатомия, брюшная полость и таз, маточные артерии Статья
Введение
Маточные артерии — это главные кровеносные сосуды, снабжающие матку кровью. Они выделяют ветви, которые снабжают различные части матки и играют важную роль в поддержании кровоснабжения во время физиологических процессов, таких как изменение эндометрия во время менструального цикла и рост матки во время беременности.По ходу маточная артерия проходит кпереди от дистального отдела мочеточника; некоторые называют это «водой под мостом».
Устройство и функции
Есть две маточные артерии, по одной с каждой стороны (левая и правая). Маточная артерия — это ветвь внутренней подвздошной артерии, также называемой гипогастральной артерией. Правая маточная артерия — это ветвь правого переднего отдела внутренней подвздошной артерии. Маточная артерия проходит от латерального к медиальному направлению через нижнюю часть широкой связки, также называемой кардинальной связкой.На уровне перешейка матки маточная артерия делится на восходящую и нисходящую ветви. [1] [2] [3]
После бифуркации восходящая артерия поднимается вверх, рядом с маткой, и проходит извилистым путем с боковой стороной матки. Восходящая артерия у повторнородящих женщин становится более извилистой и извилистой по размеру и форме. Восходящая ветвь маточной артерии анастомозирует с яичниковой артерией. Яичниковая артерия — прямая ветвь брюшной аорты.Яичниковая артерия поставляет насыщенную кислородом кровь в яичник, маточную трубу и матку. В миометрии маточная артерия разветвляется на дугообразную артерию, лучевую артерию, спиральную артерию и базальную артерию.
Нисходящая ветвь снабжает кровью шейку матки и влагалище. Дугообразная артерия вокруг шейки матки также называется круговой артерией шейки матки. Маточная артерия также ведет через нее ветви к маточной трубе и мочеточнику. Цервиковагинальные ветви анастомозируют с вагинальными артериями, образуя непроходимые артерии влагалища.
Дугообразные артерии матки снабжают кровью гладкую мускулатуру матки, называемую миометрием. Дугообразные артерии проникают по окружности миометрия и снабжают кровью переднюю и заднюю стенки матки. Дугообразная артерия заканчивается спиральной артерией, которая снабжает эндометрий, децидуальную оболочку и плаценту во время беременности. Ритмичное сокращение и расслабление этих сосудов помогает поддерживать стабильность эндометрия и необходимый контроль кровоснабжения.Эти сосуды обычно проявляются в поздней лютеиновой фазе. Перед началом менструации и во время менструации эти сосуды менее заметны. Эстроген вызывает увеличение размера и количества этих сосудов и увеличивает кровоток. Эстроген отвечает за разрастание эндометрия матки во время пролиферативной фазы менструального цикла.
Спиральные артерии снабжают кровью эндометрий матки, а точнее функциональную зону, которая отходит во время менструации.В гистологии выявление спиральных артерий помогает поставить диагноз лютеиновой фазы менструального цикла.
Базальные артерии снабжают кровью эндометрий, а точнее базальную зону. В базальной зоне начинается регенерация обнаженного эндометрия после менструации. Эта регенерация происходит под действием эстрогена во время пролиферативной фазы менструального цикла. Поддержание стромы происходит с помощью прогестерона. [4]
Хирургические рекомендации
При патологических состояниях, таких как миома матки, ход маточной артерии и ее ветвей значительно искажается.Миома матки — самая распространенная опухоль матки у женщин репродуктивного возраста. Маточная артерия снабжает кровью растущие миомы, и это кровоснабжение способствует их росту. Большинство миомы доброкачественные и бессимптомные. Симптоматические миомы могут вызывать сильную тазовую боль, дисменорею, меноррагию и железодефицитную анемию. Симптоматические миомы являются наиболее частым показанием к гистерэктомии в США. Симптоматические миомы обычно лечат хирургическим путем. После репродуктивного возраста миому матки можно лечить гонадотропин-рилизинг-гормоном (ГнРГ) для уменьшения размеров миомы.Эмболизация маточной артерии блокирует приток крови к миомам, уменьшая их в размерах; эта радиологическая процедура может улучшить симптомы у многих пациентов. Не рекомендуется в репродуктивном периоде, так как может вызвать атрофию яичников, что приведет к симптомам менопаузы. [5]
Во время гистерэктомии важно изолировать нисходящий мочеточник от маточной артерии, чтобы избежать повреждения мочеточника. Хирургическое повреждение нижней части мочеточника происходит во время сложной операции, например, при тубо-яичниковом абсцессе, образовании таза или различных злокачественных заболеваниях органов малого таза.Часто во время операции упускаются травмы мочеточника. Послеоперационная цистоскопия и визуализация свободного истечения красителя из обоих мочеточников могут гарантировать сохранность мочеточников [6].
Во время кесарева сечения может быть обильное кровотечение во время операции. Наложение швов или сдавливание матки или медицинское вмешательство могут оказаться безуспешными. В этой ситуации двусторонняя перевязка маточной артерии может остановить кровотечение. После выздоровления автоматически происходит реканализация маточных сосудов.
Клиническая значимость
Беременность — это физиологическое состояние, при котором гладкие мышцы матки и кровеносные сосуды непрерывно разрастаются.Во время беременности наблюдается значительное снижение сосудистого и мышечного тонуса матки и усиление маточного кровотока. Повышенное количество стероидных гормонов во время беременности, таких как эстроген, прогестерон и кортизол, влияет на сосудистую сеть и мускулатуру матки. Эстроген является основным фактором увеличения размера матки и утолщения стенки матки. [4]
Миометрий, средний слой стенки матки, в основном состоит из гладкомышечных клеток. Миометрий при беременности сильно увеличивается.Во время нормального гестационного периода сокращений матки не наблюдается. Предотвращение сокращения достигается за счет подавления прогестерона миоцитов. Во время беременности миоциты подвергаются постоянному растяжению, гипертрофии и ремоделированию. Эти физиологические изменения необходимы для успешной беременности. На ранних сроках беременности вневорсинчатые цитотрофобласты проникают в децидуализированный эндометрий и миометрий. Спиральные артерии трансформируются в крупные сосуды с низким сопротивлением, что происходит с постепенной потерей нормальной мышечно-эластической структуры артериальной стенки.При преэклампсии или задержке роста плода трофобластическая инвазия и дисфункция спиральной артерии сдерживают нормальные физиологические изменения.
Во время беременности допплеровское исследование маточной артерии может использоваться для сканирования кровотока в маточной артерии. Это исследование показывает степень обструкции маточной артерии. При задержке роста плода, сахарном диабете, гипертонических расстройствах и преэклампсии наблюдается нарушение кровоснабжения плаценты и плода. Цветная визуализация маточной артерии и ультразвуковое допплеровское исследование с использованием импульсной волны могут быть полезны для оценки кровотока в маточной артерии, среднего артериального давления и индекса сопротивления маточной артерии.Беременные пациентки с повышенным средним артериальным давлением, но с нормальным индексом сопротивления маточной артерии, могут рассчитывать на благоприятный исход беременности. Пациенты с повышенным средним артериальным давлением и аномальным индексом сопротивления маточной артерии могут иметь неблагоприятный исход.
Увеличение маточно-плацентарного кровотока необходимо для нормального исхода беременности. Это достигается за счет систематического роста и ремоделирования маточного кровообращения и образования плаценты. Во время беременности диаметр основной маточной артерии увеличивается примерно вдвое.Утолщение артериальной стенки при увеличении диаметра артерии отсутствует.
У женщин в постменопаузе кальцификаты могут обнаруживаться в стенках дугообразных артерий. Эти кальцинированные пятна являются эхогенными и выглядят как линейные эхо с затемнением. Это нормальный процесс старения, который может усиливаться у пациентов с диабетом. Эти кальцифицированные поражения отличаются от кальцинированных миомы.
Регулирование ремоделирования спиральной артерии матки: обзор
Гамильтон WJ, Бойд JD. Развитие плаценты человека в первые три месяца беременности. J Anat. 1960; 94: 297–328.
CAS PubMed PubMed Central Google Scholar
Ramsey EM, Houston ML, Harris JW. Взаимодействие трофобласта и материнских тканей у трех близкородственных видов приматов. Am J Obstet Gynecol. 1976; 124: 647–52.
CAS PubMed Google Scholar
Pijnenborg R, Dixon G, Robertson WB, Brosens I. Трофобластическая инвазия децидуальной оболочки человека с 8 до 18 недель беременности. Плацента. 1980; 1: 3–19.
CAS PubMed Google Scholar
Эндерс А.С., Кинг Б.Ф. Ранние стадии трофобластической инвазии сосудистой системы матери при имплантации макакам и павианам. Am J Anat. 1991; 192: 329–46.
CAS PubMed Google Scholar
Анант CV. Ишемическая плацентарная болезнь: объединяющая концепция преэклампсии, задержки внутриутробного развития и отслойки плаценты. Семин Перинатол. 2014; 38: 131–2.
PubMed Google Scholar
Heazell AE, Worton SA, Higgins LE, Ingram E, Johnstone ED, Jones RL, et al. Лекция IFPA Gábor Than Award: признание плацентарной недостаточности является ключом к спасению жизней младенцев. Плацента. 2015; 36: S20–8.
CAS PubMed Google Scholar
Brosens I. Исследование спиральных артерий базальной децидуальной оболочки при беременностях с нормальным и гипертоническим давлением. J Obstet Gynaecol Br Commonw. 1964; 71: 222–30.
CAS PubMed Google Scholar
Кхонг Т.Ю., Де Вольф Ф., Робертсон В.Б., Бросенс И. Неадекватная реакция материнских сосудов на плацентацию при беременности, осложненной преэклампсией, и у младенцев, не достигших гестационного возраста. Br J Obstet Gynaecol. 1986; 93: 1049–59.
CAS PubMed Google Scholar
Пийненборг Р., Веркрюсс Л., Хансенс М. Спиральные артерии матки при беременности человека: факты и противоречия. Плацента. 2006; 27: 939–58.
CAS PubMed Google Scholar
Lyall F, Robson SC, Bulmer JN. Ремоделирование спиральной артерии и инвазия трофобластов при преэклампсии и задержке роста плода: взаимосвязь с клиническим исходом.Гипертония. 2013; 62: 1046–54.
CAS PubMed Google Scholar
Brosens I, Puttemans P, Benagiano G. Исследование плацентарного ложа: I. Плацентарное ложе: от ремоделирования спиральных артерий до больших акушерских синдромов. Am J Obstet Gynecol. 2019; 221 (5): 437–56.
PubMed Google Scholar
Myatt L, Roberts JM. Преэклампсия: синдром или заболевание? Curr Hypertens Rep.2015; 17 (11): 83.
PubMed Google Scholar
Burton GJ, Jauniaux E. Патофизиология ограничения роста плода из плаценты. Am J Obstet Gynecol. 2018; 218: S745–61.
CAS PubMed Google Scholar
Робертс Дж. М., Тейлор Р. Н., Голдфьен А. Клинические и биохимические доказательства дисфункции эндотелиальных клеток при преэклампсии синдрома беременности.Am J Hypertens. 1991; 4: 700–8.
CAS PubMed Google Scholar
Gilbert JS, Ryan MJ, LaMarca BB, Sedeek M, Murphy SR, Granger JP. Патофизиология гипертонии во время преэклампсии: связь ишемии плаценты с эндотелиальной дисфункцией. Am J Physiol Heart Circ Physiol. 2008; 294: H541–50.
CAS PubMed Google Scholar
Робертс Дж. М., Белл М. Дж.Если мы так много знаем о преэклампсии, почему мы не вылечили болезнь? J Reprod Immunol. 2013; 99: 1–9.
PubMed PubMed Central Google Scholar
Палей А.К., Спрэдли Ф.Т., Уоррингтон, JP, Джордж Э.М., Грейнджер, JP. Патофизиология артериальной гипертензии при преэклампсии: урок интегративной физиологии. Acta Physiol. 2013; 208: 224–33.
CAS Google Scholar
Granger JP, Spradley FT, Bakrania BA. Эндотелиновая система: важнейший участник патофизиологии преэклампсии. Curr Hypertens Rep.2018; 20:32.
PubMed PubMed Central Google Scholar
Рана С., Лемуан Э, Грейнджер Дж., Каруманчи С.А. Преэклапмсия. Circ Res. 2019; 124: 1094–112.
CAS PubMed Google Scholar
Персонал AC.Двухэтапная плацентарная модель преэклампсии: обновленная информация. J Reprod Immunol. 2019; 134–135: 1–10.
PubMed Google Scholar
Тантбиройн П., Крам С.П., Параст ММ. Патофизиология крета плаценты: роль децидуальной оболочки и вневорсинчатого трофобласта. Плацента. 2008; 29: 639–45.
CAS PubMed Google Scholar
Хэннон Т., Иннес Б.А., Лэш Г.Е., Балмер Дж. Н., Робсон С.К.Влияние локальной децидуальной оболочки на инвазию трофобластов и ремоделирование спиральной артерии в очаговой крете плаценты — иммуногистохимическое исследование. Плацента. 2012; 33: 998–1004.
CAS PubMed Google Scholar
Чжоу Ю., Макмастер М., Ву К., Джанатпур М., Перри Дж., Карпанен Т. и др. Лиганды и рецепторы фактора роста эндотелия сосудов, которые регулируют выживаемость цитотрофобластов человека, не регулируются при тяжелой преэклампсии и гемолизе, повышенном уровне ферментов печени и синдроме низких тромбоцитов.Am J Pathol. 2002; 160: 1405–23.
CAS PubMed PubMed Central Google Scholar
Лала П.К., Чакраборти С. Факторы, регулирующие миграцию и инвазивность трофобластов: возможные нарушения, способствующие преэклампсии и повреждению плода. Плацента. 2003. 24: 575–87.
CAS PubMed Google Scholar
Антеби Е.Ю., Гринфилд С., Натансон-Ярон С., Гольдман-Воль Д., Хамани Ю., Худяк В. и др.Фактор роста эндотелия сосудов, фактор роста эпидермиса и факторы роста фибробластов-4 и -10 стимулируют систему активатора плазминогена трофобласта и металлопротеиназу-9. Мол Хум Репрод. 2004. 10: 229–35.
CAS PubMed Google Scholar
Leach RE, Килберн Б., Ван Дж., Лю З., Ромеро Р., Армант ДР. Гепарин-связывающий EGF-подобный фактор роста регулирует развитие вневорсинчатых цитотрофобластов человека во время перехода к инвазивному фенотипу.Dev Biol. 2004; 266: 223–37.
CAS PubMed Google Scholar
Лайалл Ф. Прайминг и ремоделирование спиральных артерий плацентарного ложа человека во время беременности — обзор. Плацента. 2005; 26: S31–6.
PubMed Google Scholar
Рао К.В., Лей З.М. Прошлое, настоящее и будущее негонадальных действий ЛГ / ХГЧ в репродуктивной биологии и медицине. Mol Cell Endocrinol.2007; 269: 2–8.
CAS PubMed Google Scholar
Harris LK. Обзор: Взаимодействие трофобластов и сосудов на ранних сроках беременности: как реконструировать сосуд. Плацента. 2010; 31: S93–8.
PubMed Google Scholar
Knöfler M, Pollheimer J. Премия IFPA в области плацентологии, лекция: молекулярная регуляция инвазии трофобластов человека. Плацента. 2012; 33: S55–62.
PubMed PubMed Central Google Scholar
Lala N, Girish GV, Cloutier-Bosworth A, Lala PK. Механизмы регуляции декорином индуцированной сосудистым эндотелиальным фактором роста миграции трофобластов человека и приобретения эндотелиального фенотипа. Биол Репрод. 2012; 87 (59): 1–14.
Google Scholar
Li Y, Zhu H, Klausen C, Peng B, Leung PC. Фактор роста эндотелия сосудов-A (VEGF-A) опосредует индуцированное активином A образование эндотелиально-подобной трубки трофобласта человека.Эндокринология. 2015; 156: 4257–68.
CAS PubMed Google Scholar
Эванс Дж., Саламонсен Л.А., Менкхорст Э., Димитрадис Э. Динамические изменения гипергликозилированного хорионического гонадотропина человека в течение первого триместра беременности и его роль в ранней плацентации. Hum Reprod. 2015; 30: 1029–38.
CAS PubMed Google Scholar
Ло Цюй, Лю Х, Чжэн И, Чжао И, Чжу Дж, Цзоу Л.Эфрин-B2 опосредует трофобласт-зависимое ремоделирование спиральной артерии матери в первом триместре. Плацента. 2015; 36: 567–74.
CAS PubMed Google Scholar
Дефорж М., Харрис Л.К., Альпин Д.Д. Пептиды, производные эластина, стимулируют миграцию и инвазию трофобластов: петля положительной обратной связи для усиления ремоделирования спиральной артерии. Мол Хум Репрод. 2015; 21: 95–104.
Google Scholar
Fan M, Xu Y, Hong F, Gao X, Xin G, Hong H и др. Путь передачи сигналов Rac1 / β-catenin способствует инвазии клеток трофобласта, нацеливаясь на улитку и MMP9. Cell Physiol Biochem. 2016; 38: 1319–32.
CAS PubMed Google Scholar
Zhao WX, Zhuang X, Huang TT, Feng R, Lin JH. Влияние Notch3 и Notch4 на пролиферацию и апоптоз клеточных линий трофобластов. Int J Med Sci. 2015; 12: 867–74.
CAS PubMed PubMed Central Google Scholar
McNally R, Alqudah A, Obradovic D, McClements L. Выяснение патогенеза преэклампсии с использованием моделей ремоделирования спиральной маточной артерии in vitro. Curr Hypertens Rep.2017; 19: 93.
PubMed PubMed Central Google Scholar
Athanassiades A, Hamilton GS, Lala PK. Фактор роста эндотелия сосудов стимулирует пролиферацию, но не миграцию или инвазивность вневорсинчатого трофобласта человека. Биол Репрод.1998. 59: 643–54.
CAS PubMed Google Scholar
Фитцпатрик Т.Э., Лэш Г.Е., Янаихара А., Чарнок-Джонс Д.С., Макдональд-Гудфеллоу С.К., Грэм Ч. Ингибирование карциномы молочной железы и инвазивности клеток трофобласта фактором роста эндотелия сосудов. Exp Cell Res. 2003. 283: 247–55.
CAS PubMed Google Scholar
Dubinsky V, Poehlmann TG, Suman P, Gentile T., Markert UR, Gutierrez G.Роль регуляторных и ангиогенных цитокинов в инвазии трофобластических клеток. Am J Reprod Immunol. 2010; 63: 193–9.
CAS PubMed Google Scholar
Форбс К., Вествуд М. Регуляция материнским фактором роста развития плаценты человека и роста плода. J Endocrinol. 2010; 207: 1–16.
CAS PubMed Google Scholar
Джонс Р.Л., Финдли Дж.К., Фарнворт П.Г., Робертсон Д.М., Уоллес Э., Саламонсен Л.А.Активин А и ингибин А по-разному регулируют металлоптотеиназы маточного матрикса человека: потенциальные взаимодействия во время децидуализации и инвазии трофобластов. Эндокринология. 2006; 147: 724–32.
CAS PubMed Google Scholar
Hu XQ, Zhang L. МикроРНК при дисфункции маточно-плацентарных сосудов. Ячейки. 2019; 8 (11): 1344.
CAS PubMed Central Google Scholar
Hoffman P, Saoudi Y, Benharouga M, Graham CH, Schaal JP, Mazouni C и др. Роль EG-VEGF в плацентации человека: физиологические и патологические последствия. J Cell Mol Med. 2009; 13: 2224–35.
Google Scholar
Tamaru S, Mizuno Y, Tochigi H, Kajihara T., Okazaki Y, Okagaki R, et al. MicroRNA-135b подавляет инвазию клеток HTR-7 / SVneo, происходящих из вневорсинчатого трофобласта, путем прямого подавления регуляции CXCL12 в условиях низкого содержания кислорода.Biochem Biophys Res Commun. 2015; 461: 421–6.
CAS PubMed Google Scholar
Pan Q, Niu HY, Cheng LF, Li XQ, Zhang QG, Ning Y. Инвазия клеточных линий трофобластов ингибируется miR-93 через MMP-2. Плацента. 2017; 53: 48–53.
CAS PubMed Google Scholar
Caniggia I, Grisaru-Gravnosky S, Kuliszewsky M, Post M, Lye SJ. Ингибирование TFG-бета 3 восстанавливает инвазивную способность вневорсинчатых трофобластов при преэкламптической беременности.J Clin Invest. 1999; 103: 1641–50.
CAS PubMed PubMed Central Google Scholar
Majali-Martinez A, Velicky P, Pollheimer J, Knöfler M, Yung HW, Burton GJ, et al. Эндотелин-1 подавляет экспрессию матриксных металлопротеиназ 14 и 15 в трофобластах человека в первом триместре через рецептор эндотелина типа B. Hum Reprod. 2017; 32: 46–54.
CAS PubMed Google Scholar
Lash GE, Pitman H, Morgan HL, Innes BA, Agwu CN, Bulmer JN. Децидуальные макрофаги: ключевые регуляторы ремоделирования сосудов при беременности человека. J Leukoc Biol. 2016; 100: 315–25.
CAS PubMed Google Scholar
Смит С.Д., Данк К.Э., Аплин Д.Д., Харрис Л.К., Джонс Р.Л. Доказательства участия иммунных клеток в ремоделировании децидуальных спиральных артериол на ранних сроках беременности человека. Am J Pathol. 2009; 174: 1959–71.
CAS PubMed PubMed Central Google Scholar
Робсон А., Харрис Л.К., Иннес Б.А., Лэш Г.Е., Альджунаиди М.М., Аплин Д.Д. и др. Естественные клетки-киллеры матки инициируют ремоделирование спиральной артерии при беременности человека. FASEB J. 2012; 26: 4876–85.
CAS PubMed Google Scholar
Choudhury RH, Dunk CE, Lye SJ, Aplin JD, Harris LK, Jones RL. Перекрестное взаимодействие вневорсинчатых трофобластов и эндотелиальных клеток опосредует инфильтрацию лейкоцитов к раннему ремоделированию стенки децидуальной спиральной артериолы.J Immunol. 2017; 198 (10): 4115–28.
CAS PubMed Google Scholar
Чжан Дж., Dunk CE, Lye SJ. Передача сигналов сфингозина регулирует ангиогенный фенотип децидуальных NK-клеток и миграцию трофобластов. Hum Reprod. 2013; 28: 3026–37.
CAS PubMed Google Scholar
Pollheimer J, Vondra S, Baltayeva J, Beristain AG, Knöfler M. Регулирование вневорсинчатых трофобластов плаценты средой матки матери.Фронт Иммунол. 2018; 9: 1–18.
Google Scholar
Мозер Г., Виндспергер К., Поллхеймер Дж., Де Соуза Лопес С.К., Хуппертц Б. Вторжение человеческого трофобласта: новые и неожиданные маршруты и функции. Histochem Cell Biol. 2018; 150: 361–70.
CAS PubMed PubMed Central Google Scholar
Harris LK, Benagiano M, D’Elios MM, Brosens I, Benagiano G. Исследование плацентарного ложа: 2.Функциональные иммунологические исследования плацентарного ложа. Am J Obstet Gynecol. 2019; 221 (5): 457–69.
CAS PubMed Google Scholar
Картер А.М. Животные модели плацентации человека — обзор. Плацента. 2007; 28 (Приложение A): S41–7.
PubMed Google Scholar
Моффет А., Лок С. Иммунология плацентации у человеческих млекопитающих. Nat Rev Immunol.2006; 6: 584–94.
Google Scholar
Бейкер П.Н., Краснов Дж., Робертс Дж. М., Йео К. Т.. Повышенные сывороточные уровни фактора роста эндотелия сосудов у пациентов с преэклампсией. Obstet Gynecol. 1995; 86: 815–21.
CAS PubMed Google Scholar
Купер Дж.С., Шарки А.М., Чарнок-Джонс Д.С., Палмер К.Р., Смит С.К. Уровни мРНК VEGF в плаценте от беременностей, осложненных преэклампсией.Br J Obstet Gynaecol. 1996. 103: 1191–6.
CAS PubMed Google Scholar
Шарки А.М., Купер Дж. С., Балмфорт Дж. Р., Макларен Дж., Кларк Д. Э., Чарнок-Джонс Д. С. и др. Уровни фактора роста эндотелия сосудов в плазме крови матери при нормальных беременностях и беременностях, осложненных преэклампсией. Eur J Clin Investig. 1996; 26: 1182–5.
CAS Google Scholar
Lyall F, Greer IA, Boswell F, Fleming R. Подавление иммунореактивности сывороточного фактора роста эндотелия сосудов при нормальной беременности и при преэклампсии. Br J Obstet Gynaecol. 1997. 104: 223–8.
CAS PubMed Google Scholar
Chung JY, Song Y, Wang Y, Magness RR, Zheng J. Дифференциальная экспрессия фактора роста эндотелия сосудов (VEGF), рецепторов VEGF, производных эндокринной железы, и рецепторов VEGF в человеческих плацентах от нормальных и преэкламптических беременностей.J Clin Endocrinol Metab. 2004. 89: 2484–90.
CAS PubMed PubMed Central Google Scholar
Гамильтон Г.С., Лисиак Дж.Дж., Хан В.К., Лала П.К. Аутокринно-паракринная регуляция инвазивности трофобластов человека с помощью инсулиноподобного фактора роста (IGF) -II и IGF-связывающего белка (IGFBP) -1. Exp Cell Res. 1998. 244: 147–56.
CAS PubMed Google Scholar
Armant DR, Fritz R, Kilburn BA, Kim YM, Nien JK, Maihle NJ, et al. Снижение экспрессии сигнальной системы эпидермального фактора роста при преэклампсии. Плацента. 2015; 6: 270–8.
Google Scholar
Fragkiadaki P, Soulitzis N, Sifakis S, Koutroulakis D, Gourvas V, Vrachnis N, et al. Подавление сигнального пути notch при поздних недоношенных и доношенных плацентах от беременностей, осложненных преэклампсией. PLoS One. 2015; 10: e0126163.
PubMed PubMed Central Google Scholar
Muñoz-Hernandez R, Miranda ML, Stiefel P, Lin RZ, Praena-Fernández JM, Dominguez-Simeon MJ, et al. Снижение уровня циркулирующих в пуповинной крови эндотелиальных колониеобразующих клеток при преэклампсии. Гипертония. 2014; 64: 165–71.
PubMed PubMed Central Google Scholar
Travaglino A, Raffone A, Saccone G, Migliorini S, Maruotti GM, Esposito G, et al.Морфология плаценты, апоптоз, ангиогенез и эпителиальные механизмы при преэклампсии с ранним началом. Eur J Obstet Gynecol Reprod Biol. 2019; 234: 200–6.
CAS PubMed Google Scholar
Chen J, Yue C, Xu J, Zhan Y, Zhao H, Li Y, et al. Подавление рецепторной тирозинкиназоподобного орфанного рецептора 1 в преэклампсии плаценты ингибирует пролиферацию, миграцию и инвазию трофобластных клеток человека посредством аккомодации пути PI3K / AKT / mTOR.Плацента. 2019; 82: 17–24.
CAS PubMed Google Scholar
Torry DS, Wang HS, Wang TH, Caudle MR, Torry RJ. Преэклампсия связана со снижением сывороточного уровня фактора роста плаценты. Am J Obstet Gynecol. 1998. 179: 1539–44.
CAS PubMed Google Scholar
Тейлор Р.Н., Гримвуд Дж., Тейлор Р.С., Макмастер М.Т., Фишер С.Дж., Север РА. Продольные сывороточные концентрации фактора роста плаценты: доказательства аномального ангиогенеза плаценты при патологических беременностях.Am J Obstet Gynecol. 2003. 188: 177–82.
CAS PubMed Google Scholar
Цатсарис В., Гоффин Ф., Мунаут С., Бричан Дж. Ф., Пиньон М. Р., Ноэль А. и др. Сверхэкспрессия рецептора растворимого сосудистого эндотелиального фактора роста у пациентов с преэклампсией: патофизиологические последствия. J Clin Endocrinol Metab. 2003. 88: 5555–63.
CAS PubMed Google Scholar
Maynard SE, Min JY, Merchan J, Lim KH, Li J, Mondal S и др. Избыток плацентарной растворимой fms-подобной тирозинкиназы 1 (sFlt1) может способствовать эндотелиальной дисфункции, гипертонии и протеинурии при преэклампсии. J Clin Invest. 2003. 111: 649–58.
CAS PubMed PubMed Central Google Scholar
Levine RJ, Maynard SE, Qian C, Lim KH, England LJ, Yu KF, et al. Циркулирующие ангиогенные факторы и риск преэклампсии.N Engl J Med. 2004; 350: 672–83.
CAS PubMed Google Scholar
Чайворапонгса Т., Ромеро Р., Ким Ю.М., Ким Г.Дж., Ким М.Р., Эспиноза Дж. И др. Концентрация растворимого в плазме рецептора-1 фактора роста эндотелия сосудов повышается до постановки клинического диагноза преэклампсии. J Matern Fetal Neonatal Med. 2005; 17: 3–18.
CAS PubMed Google Scholar
Wathén KA, Tuutti E, Stenman UH, Alfthan H, Halmesmäki E, Finne P, et al. Рецептор-1 растворимого в сыворотке крови фактора роста эндотелия сосудов матери на ранних сроках беременности, заканчивающейся преэклампсией или задержкой внутриутробного развития. J Clin Endocrinol Metab. 2006; 91: 180–4.
PubMed Google Scholar
Левин Р.Дж., Лам К., Цянь С., Ю К.Ф., Мейнард С.Е., Сакс Б.П. и др. Растворимый эндоглин и другие циркулирующие антиангиогенные факторы при преэклампсии.N Engl J Med. 2006; 355: 992–1005.
CAS PubMed Google Scholar
Myatt L, Clifton RG, Roberts JM, Spong CY, Hauth JC, Varner MW и др. Прогнозирование преэклампсии в первом триместре у первородящих женщин с низким риском. Obstet Gynecol. 2012; 119: 1234–42.
PubMed PubMed Central Google Scholar
Kenny LC, Black MA, Poston L, Taylor R, Myers JE, Baker PN, et al.Раннее прогнозирование преэклампсии у первородящих женщин с учетом клинического риска и биомаркеров: международное когортное исследование «Скрининг конечных точек беременности» (SCOPE). Гипертония. 2014; 64: 644–52.
CAS PubMed Google Scholar
Серин И.С., Кула М., Башбуг М., Унлухизарчи К., Гучер С., Тайяр М. Уровни андрогенов у пациенток с преэклампсией в третьем триместре беременности и через шесть недель после родов. Acta Obstet Gynecol Scand.2001; 80: 1009–13.
CAS PubMed Google Scholar
Zeisler H, Jirecek S, Hohlagschwandtner M, Knöfler M, Tempfer C, Livingston JC. Концентрации эстрогенов у пациентов с преэклампсией. Wien Klin Wochenschr. 2002; 114: 458–61.
CAS PubMed Google Scholar
Миллер Н.Р., Гарри Д., Коэн Х.В., Фигероа Р. Сывороточные маркеры андрогенов при преэклампсии.J Reprod Med. 2003. 48: 225–9.
PubMed Google Scholar
Salas SP, Marshall G, Gutiérrez BL, Rosso P. Динамика объема материнской плазмы и гормональных изменений у женщин с преэклампсией или задержкой роста плода. Гипертония. 2006; 47: 203–8.
CAS PubMed Google Scholar
Hertig A, Liere P, Chabbert-Buffet N, Fort J, Pianos A, Eychenne B, et al.Профилирование стероидов у женщин с преэклампсией: доказательства дефицита ароматазы. Am J Obstet Gynecol. 2010; 203: 477.e1–9.
Google Scholar
Berkane N, Liere P, Oudinet JP, Hertig A, Lefèvre G, Pluchino N, et al. От беременности до преэклампсии: ключевая роль эстрогенов. Endocr Rev.2017; 38: 123–44.
PubMed Google Scholar
Джоб С.О., Тайлер К.Т., Магнесс Р.Р.Аберрантный синтез, метаболизм и накопление в плазме циркулирующих эстрогенов и метаболитов эстрогенов в последствиях преэклампсии для сосудистой дисфункции. Гипертония. 2013; 61: 480–7.
CAS PubMed PubMed Central Google Scholar
Faas MM, De Vos P. Врожденные иммунные клетки в плацентарном ложе при здоровой беременности и преэклампсии. Плацента. 2018; 29: 125–33.
Google Scholar
Stallmach T, Hebisch G, Orban P, Lu X. Аберрантное расположение трофобластов и лимфоцитов на стыке плода и матери с преэклампсией. Арка Вирхова. 1999; 434 (3): 07–211.
Google Scholar
Bachmayer N, Rafik Hamad R, Liszka L, Bremme K, Sverremark-Ekstrom E. Аберрантная экспрессия маточных естественных киллеров (NK) и измененные уровни NK-клеток в плаценте и сыворотке, способствующие цитокину интерлейкина-12 при преэклампсии.Am J Reprod Immunol. 2006. 56 (5–6): 292–301.
CAS PubMed Google Scholar
Wilczyński JR, Tchórzewski H, Banasik M, Głowacka E, Wieczorek A, Lewkowicz P, et al. Распределение субпопуляций лимфоцитов и секреция цитокинов в децидуальной оболочке третьего триместра при нормальной беременности и преэклампсии. Eur J Obstet Gynecol Reprod Biol. 2003. 109 (1): 8–15.
PubMed Google Scholar
Schonkeren D, van der Hoorn ML, Khedoe P, Swings G, van Beelen E, Claas F и др. Дифференциальное распределение и фенотип децидуальных макрофагов при преэкламптических беременностях по сравнению с контрольными. Am J Pathol. 2011. 178 (2): 709–17.
CAS PubMed PubMed Central Google Scholar
Рейстер Ф., Франк Х.Г., Королевство Дж. К., Хейл В., Кауфманн П., Рат В. и др. Апоптоз, индуцированный макрофагами, ограничивает эндоваскулярную инвазию трофобластов в стенку матки у женщин с преэклампсией.Lab Investig. 2001. 81 (8): 1143–52.
CAS PubMed Google Scholar
Kim JS, Romero R, Cushenberry E, Kim YM, Erez O, Nien JK, et al. Распределение макрофагов CD14 + и CD68 + в плацентарном ложе и базальной пластине у женщин с преэклампсией и преждевременными родами. Плацента. 2007. 28 (5–6): 571–6.
CAS PubMed Google Scholar
Уильямс П.Дж., Балмер Дж. Н., Сирл Р.Ф., Иннес Б.А., Робсон СК.Измененные популяции децидуальных лейкоцитов в плацентарном ложе при преэклампсии и задержке роста плода: сравнение с нормальной беременностью на поздних сроках. Репродукция. 2009. 138 (1): 177–84.
CAS PubMed Google Scholar
Eide IP, Rolfseng T, Isaksen CV, Mecsei R, Roald B., Lydersen S, et al. Серьезное ограничение роста плода связано с уменьшением доли естественных клеток-киллеров в децидуальной оболочке. Арка Вирхова.2006. 448 (3): 269–76.
PubMed Google Scholar
Bürk MR, Troeger C, Brinkhaus R, Holzgreve W., Hahn S. Сильно уменьшенное присутствие тканевых макрофагов в базальной пластинке преэкламптических плацент. Плацента. 2001. 22 (4): 309–16.
PubMed Google Scholar
Ригер Л., Сегерер С., Бернар Т., Капп М., Маджич М., Морр А.К. и др. Конкретные подмножества иммунных клеток децидуальной оболочки человека различаются при нормальной беременности и преэклампсии — проспективное обсервационное исследование.Reprod Bio Endocrino: RBE (Rev Bras Entomol). 2009; 7: 132.
Google Scholar
Хсу П., Сантнер-Нанан Б., Дальстром Дж. Э., Фадиа М., Чандра А., Пик М. и др. Измененные децидуальные антигенпрезентирующие клетки DC-SIGN + и нарушение индукции регуляторных Т-клеток при преэклампсии. Am J Pathol. 2012. 181 (6): 2149–60.
CAS PubMed Google Scholar
Хуанг С.Дж., Чен С.П., Шатц Ф., Рахман М., Абрахамс В.М., Локвуд С.Дж.Преэклампсия связана с привлечением дендритных клеток в децидуальную оболочку матки. J Pathol. 2008. 214 (3): 328–36.
CAS PubMed Google Scholar
Hiby SE, Walker JJ, O’Shaughnessy KM, Redman CW, Carrington M, Trowsdale J, et al. Комбинации материнских генов KIR и HLA-C плода влияют на риск преэклампсии и репродуктивный успех. J Exp Med. 2004. 200 (8): 957–65.
CAS PubMed PubMed Central Google Scholar
Рейстер Ф., Франк Х. Г., Хейл В., Косанке Г., Хуппертц Б., Шредер В. и др. Распределение макрофагов в спиральных артериях плацентарного ложа при преклампсии отличается от такового у здоровых пациентов. Плацента. 1999. 20 ((2–3)): 229–33.
CAS PubMed Google Scholar
Катабучи Х., Йих С., Охба Т., Мацуи К., Такахаши К., Такея М. и др. Характеристика макрофагов в децидуальной атеротической спиральной артерии с особым акцентом на цитологию пенистых клеток.Med Electron Microsc. 2003. 36 (4): 253–62.
PubMed Google Scholar
Haeger M, Unander M, Norder-Hansson B., Tylman M, Bengtsson A. Активация комплемента, нейтрофилов и макрофагов у женщин с тяжелой преэклампсией и синдромом гемолиза, повышенными ферментами печени и низким количеством тромбоцитов. Obstet Gynecol. 1992. 79 (1): 19–26.
CAS PubMed Google Scholar
Benyo DF, Smarason A, Redman CW, Sims C, Conrad KP. Экспрессия воспалительных цитокинов в плаценте женщин с преэклампсией. J Clin Endocrinol Metab. 2001. 86 (6): 2505–12.
CAS PubMed Google Scholar
Wang Y, Walsh SW. Концентрация TNF-альфа и экспрессия мРНК увеличиваются в преэкламптической плаценте. J Reprod Immunol. 1996. 32 (2): 157–69.
CAS PubMed Google Scholar
Pang ZJ, Xing FQ. Сравнительное исследование экспрессии генов цитокинов-рецепторов в нормальной и преэкламптической плаценте человека с использованием ДНК-микрочипов. J Perinat Med. 2003. 31 (2): 153–62.
CAS PubMed Google Scholar
Хеннесси А., Пилмор Х.Л., Симмонс Л.А., Художник Д.М. Дефицит плацентарного ИЛ-10 при преэклампсии. J Immunol. 1999. 163 (6): 3491–5.
CAS PubMed Google Scholar
Rein DT, Breidenbach M, Hönscheid B, Friebe-Hoffmann U, Engel H, Göhring UJ, et al. У женщин с преэклампсией наблюдается дефицит интерлейкина-10, о чем свидетельствует высвобождение цитокинов клетками трофобласта in vitro. Цитокин. 2003. 23 (4–5): 119–25.
CAS PubMed Google Scholar
Prins JR, Faas MM, Melgert BN, Huitema S, Timmer A, Hylkema MN, et al. Измененная экспрессия иммуноассоциированных генов в децидуальной оболочке человека в первом триместре беременностей, позже осложненных гипертонией или задержкой роста плода.Плацента. 2012; 33 (5): 453–5.
CAS PubMed Google Scholar
Редман CW, Сарджент Иллинойс, персонал AC. Лекция IFPA Senior Award: понимание преэклампсии — две плацентарные причины преэклампсии? Плацента. 2014; 35 (Прил.): S20–5.
PubMed Google Scholar
Робертс Дж. М., Тейлор Р. Н., Муски Т. Дж., Роджерс Дж. М., Хьюбел, Калифорния, Маклафлин М.К. Преэклампсия: нарушение эндотелиальных клеток.Am J Obstet Gynecol. 1989. 161: 1200–4.
CAS PubMed Google Scholar
Барден А., Бейлин Л.Дж., Ричи Дж., Уолтерс Б.Н., Майкл, Калифорния. Плазма и мочевой эндотелин 1, метаболиты простациклина и потребление тромбоцитов при преэклампсии и гипертонической беременности. Кровавый пресс. 1994; 3: 38–46.
CAS PubMed Google Scholar
Уолш SW.Материнско-плацентарное взаимодействие оксидативного стресса и антиоксидантов при преэклампсии. Семин Репрод Эндокринол. 1998. 16: 93–104.
CAS PubMed Google Scholar
Мятт Л., Вебстер Р.П. Сосудистая биология преэклампсии. J Thromb Haemost. 2009. 7: 375–84.
CAS PubMed Google Scholar
Каннингем Ф.Г., Робертс Дж. М., Линдхаймер Мэриленд. Клинический спектр преэклампсии.В: Lindheimer MD, Roberts JM, Cunningham FG, редакторы. Гипертоническая болезнь Чесли при беременности. Сан-Диего: академический; 2009. с. 25–36.
Google Scholar
Редман CW, Сарджент Иллинойс. Плацентарный стресс и преэклампсия: новый взгляд. Плацента. 2009; 30 (Приложение A): S38–42.
PubMed Google Scholar
Brennan LJ, Morton JS, Davidge ST. Сосудистая дисфункция при преэклампсии.Микроциркуляция. 2014; 21: 4–14.
PubMed Google Scholar
Goulopoulou S, Davidge ST. Молекулярные механизмы материнской сосудистой дисфункции при преэклампсии. Тенденции Мол Мед. 2015; 21: 88–97.
CAS PubMed Google Scholar
Халил Р.А., Грейнджер Дж.П. Сосудистые механизмы повышения артериального давления при преэклампсии: уроки на животных моделях.Am J Physiol Regul Integr Comp Physiol. 2002; 283: R29–45.
CAS PubMed Google Scholar
Ilekis JV, Tsilou E, Fisher S, Abrahams VM, Soares MJ, Cross JC, et al. Плацентарное происхождение неблагоприятных исходов беременности: потенциальные молекулярные мишени: Резюме семинара для руководителей Национального института здоровья детей и развития человека Юнис Кеннеди Шрайвер. Am J Obstet Gynecol. 2016; 215: S1 – S46.
PubMed PubMed Central Google Scholar
Sladek SM, Magness RR, Conrad KP. Оксид азота и беременность. Am J Phys. 1997; 272: R441–63.
CAS Google Scholar
Худ Дж. Д., Мейнингер Си Джей, Зиче М., Грейнджер Х. Дж. VEGF активизирует сигнал ecNOS, выработку белка и NO в эндотелиальных клетках человека. Am J Phys. 1998; 274: h2054–8.
CAS Google Scholar
Ferrara N, Gerber HP, LeCouter J.Биология VEGF и его рецепторов. Nat Med. 2003. 9: 669–76.
CAS Google Scholar
Венкатеша С., Топорсиан М., Лам К., Ханаи Дж., Маммото Т., Ким Ю.М. и др. Растворимый эндоглин вносит свой вклад в патогенез преэклампсии. Nat Med. 2006; 12 (6): 642–9 Epub 2006 4 июня.
CAS PubMed Google Scholar
Лу Ф., Лонго М., Тамайо Э., Манер В., Аль-Хенди А., Андерсон Г.Д. и др.Влияние сверхэкспрессии sFlt-1 на кровяное давление и возникновение других проявлений преэклампсии у несдержанных беременных мышей в сознании. Am J Obstet Gynecol. 2007; 196: 396.e1–7.
Google Scholar
Tang JR, Karumanchi SA, Seedorf G, Markham N, Abman SH. Избыток растворимого рецептора-1 фактора роста эндотелия сосудов в околоплодных водах ухудшает рост легких у крыс: связывая преэклампсию с бронхолегочной дисплазией.Am J Physiol Lung Cell Mol Physiol. 2012; 302: L36–46.
CAS PubMed Google Scholar
LaMarca BD, Gilbert JS, Granger JP. Недавний прогресс в понимании патофизиологии гипертонии во время преэклампсии. Гипертония. 2008. 51: 982–8.
CAS PubMed PubMed Central Google Scholar
Suzuki H, Ohkuchi A, Matsubara S, Takei Y, Murakami M, Shibuyua M и др.Влияние рекомбинантного фактора роста плаценты 2 на гипертензию, индуцированную полноразмерным мышиным растворимым аденовирусным вектором fms-подобной тирозинкиназы 1 у беременных мышей. Гипертония. 2009; 54: 112
.Google Scholar
Bridges JP, Gilbert JS, Colson D, Gilbert SA, Dukes MP, Ryan MJ, et al. Окислительный стресс способствует индуцированной растворимой fms-подобной тирозинкиназе-1 сосудистой дисфункции у беременных крыс. Am J Hypertens. 2009; 22: 564–8 Epub 2009 5 марта.
CAS PubMed PubMed Central Google Scholar
Szalai G, Romero R, Chaiworapongsa T, Xu Y, Wang B, Ahn H, et al. Полноразмерная изоформа sFlt-1-e15a плаценты человека индуцирует различные материнские фенотипы преэклампсии у мышей. PLoS One. 2015; 10 (4): e0119547.
PubMed PubMed Central Google Scholar
Ханкин Э.В., Мандала М, Колтон I, Каруманчи С.А., Осол Г.Гемодинамическое, сосудистое и репродуктивное влияние блокады FMS-подобной тирозинкиназы 1 (FLT1) на маточно-плацентарное кровообращение во время нормальной беременности мышей. Биол Репрод. 2012; 86: 57,1–8.
Google Scholar
Li Z, Zhang Y, Ying Ma J, Kapoun AM, Shao Q, Kerr I., et al. Рекомбинантный фактор роста эндотелия сосудов 121 ослабляет гипертензию и улучшает повреждение почек на модели преэклампсии у крыс. Гипертония. 2007; 50: 686–92.
CAS PubMed Google Scholar
Бергманн А., Ахмад С., Кадмор М., Грубер А.Д., Витчен П., Линденмайер В. и др. Уменьшение циркулирующего растворимого Flt-1 облегчает симптомы, подобные преэклампсии, у мышей. J Cell Mol Med. 2010. 14: 1857–67.
CAS PubMed Google Scholar
Gilbert JS, Verzwyvelt J, Colson D, Arany M, Karumanchi SA, Granger JP.Инфузия рекомбинантного фактора роста эндотелия сосудов 121 снижает кровяное давление и улучшает функцию почек у крыс с гипертензией, вызванной ишемией плаценты. Гипертония. 2010. 55: 380–385.
Woods AK, Hoffmann DS, Weydert CJ, Butler SD, Zhou Y, Sharma RV и др. Аденовирусная доставка VEGF121 на ранних сроках беременности предотвращает спонтанное развитие преэклампсии у мышей с ДГПЖ / 5. Гипертония. 2011; 57: 94–102.
CAS PubMed Google Scholar
Mateus J, Bytautiene E, Lu F, Tamayo EH, Betancourt A, Hankins GD, et al. Терапия эндотелиальным фактором роста улучшает проявления, подобные преэклампсии, на мышиной модели, индуцированные сверхэкспрессией sVEGFR-1. Am J Physiol Heart Circ Physiol. 2011; 301: h2781–7.
CAS PubMed Google Scholar
Карр Д. Д., Уоллес Дж. М., Эйткен Р. П., Милн Дж. С., Мартин Дж. Ф., Захари И. К. и др. Пери- и постнатальные эффекты пренатальной аденовирусной генной терапии VEGF у овец с ограниченным ростом.Биол Репрод. 2016; 94: 142,1–12.
Google Scholar
Кумасава К., Икава М., Кидоя Х., Хасува Х., Сайто-Фудзита Т., Мориока Ю. и др. Правастатин индуцирует фактор роста плаценты (PGF) и уменьшает преэклампсию на модели мышей. Proc Natl Acad Sci U S. A. 2011; 108: 1451–5.
CAS PubMed Google Scholar
Rejinders D, Liu CC, Xu X, Zhao AM, Olson KN, Butler SD, et al.Целекоксиб восстанавливает экспрессию ангиогенного фактора на границе раздела матери и плода в модели преэклампсии на мышах с ДГПЖ / 5. Physiol Genomics. 2018; 50 (5): 385–92.
Google Scholar
Ааса К.Л., Заван Б., Луна Р.Л., Вонг П.Г., Вентура Н.М., Це М.Ю. и др. Фактор роста плаценты влияет на адаптацию материнской сердечно-сосудистой системы к беременности у мышей. Биол Репрод. 2015; 92: 44.
PubMed Google Scholar
Кей В.Р., Рэтсеп М.Т., Фигейро-Филью Э.А., Крой Б.А. Преэклампсия может влиять на нейроанатомию и когнитивные функции потомства: роль фактора роста плаценты. Биол Репрод. 2019; 101: 271–83.
PubMed Google Scholar
ЛаМарка ББ, Беннетт Вашингтон, Александр БТ, Кокрелл К., Грейнджер ДжП. Гипертония, вызванная снижением перфузии матки у беременных крыс: роль фактора некроза опухоли-α. Гипертония. 2005; 46: 1022–5.
CAS PubMed Google Scholar
Gilbert JS, Babcock SA, Granger JP. Гипертония, вызванная снижением перфузии матки у беременных крыс, связана с повышенной экспрессией растворимой fms-подобной тирозинкиназы-1. Гипертония. 2007; 50: 1142–7 Epub 2007 8 октября.
CAS PubMed Google Scholar
Уолш С.К., Инглиш Ф.А., Джонс Э.Дж., Кенни Л.К.Плазма-опосредованная дисфункция сосудов в модели преэклампсии с пониженным перфузионным давлением матки: характеристика микрососудов. Гипертония. 2008; 54: 345–51.
Google Scholar
Лин, К., Хе Х, Цуй Н, Рен З, Чжу М., Халил Р.А. Снижение васкуляризации матки и расширяющееся ремоделирование маточных артерий с пониженным содержанием матриксной металлопротеиназы-2 и -9 при гипертонической беременности. Am J Physiol Heart Circ Physiol. 2019; 318 (1): h265 – h280.
Молнар М., Сютё Т., Тот Т., Хертеленди Ф. Длительная блокада синтеза оксида азота у беременных крыс вызывает стойкую гипертензию, протеинурию, тромбоцитопению и задержку внутриутробного развития плода. Am J Obstet Gynecol. 1994; 170: 1458–66.
PubMed Google Scholar
Виншип А.Л., Кога К., Менкхорст Э., Ван Синдерен М., Райнчук К., Нагай М. и др. Интерлейкин-11 изменяет плаценту и вызывает признаки преэклампсии у мышей.Proc Natl Acad Sci U S. A. 2015; 112: 15928–33.
CAS PubMed PubMed Central Google Scholar
Куландавелу С., Уайтли К.Дж., Ку Д., Мю Дж., Бейнбридж С.А., Адамсон С.Л. Дефицит эндотелиальной синтазы оксида азота снижает кровоток в матке, удлинение спиральной артерии и оксигенацию плаценты у беременных мышей. Гипертония. 2012; 60: 231–8.
CAS PubMed Google Scholar
Cureton N, Kortokova I, Baker B, Greenwood S, Wareing M, Kotamraju VR, et al. Селективное нацеливание нового вазодилататора на сосудистую сеть матки для лечения нарушения маточно-плацентарной перфузии во время беременности. Теронастика. 2017; 7 (15): 3715–31 eCollection 2017.
CAS. Google Scholar
Hunkapillar NM, Gasperowicz M, Kapidzic M, Plaks V, Maltepe E, Kitajewski J, et al. Роль передачи сигналов Notch в эндоваскулярной инвазии трофобластов и в патогенезе преэклампсии.Разработка. 2011; 138: 2987–98.
Google Scholar
Doridot L, Passet B, Méhats C, Rigourd V, Barbaux S, Ducat A, et al. Симптомы, подобные преэклампсии, вызванные у мышей фетоплацентарной экспрессией STOX1, устраняются лечением аспирином. Гипертония. 2013; 61: 662–8.
CAS PubMed Google Scholar
Гринвуд Дж. Д., Минхас К., ди Санто Дж. П., Макита М., Кисо Й., Крой Б.А.Ультраструктурные исследования участков имплантации от мышей с дефицитом естественных клеток-киллеров матки. Плацента. 2000; 21: 693–702.
CAS PubMed Google Scholar
Ашкар А.А., Ди Санто Дж. П., Крой Б.А. Гамма-интерферон способствует инициации модификации сосудов матки, децидуальной целостности и созреванию естественных клеток-киллеров матки во время нормальной беременности мышей. J Exp Med. 2000; 192: 259–70.
CAS PubMed PubMed Central Google Scholar
Крой Б.А., Хе Х., Эсадег С., Вэй К., Маккартни Д., Чжан Дж. И др. Маточные естественные клетки-киллеры: понимание их клеточной и молекулярной биологии на основе моделирования на мышах. Репродукция. 2003; 126: 149–60.
CAS PubMed PubMed Central Google Scholar
Монк Дж. М., Леонард С., Макби Б. А., Крой Б. А.. Индукция модификации спиральной артерии мыши рекомбинантным человеческим гамма-интерфероном. Плацента. 2005; 26: 835–8.
CAS PubMed Google Scholar
Надкарни С., Смит Дж., Сферруцци-Перри А.Н., Ледвозыв А., Кишор М., Хаас Р. и др. Нейтрофилы индуцируют проангиогенные Т-клетки с регуляторным фенотипом во время беременности. Proc Natl Acad Sci U S. A. 2016; 113: E8415–24.
CAS PubMed PubMed Central Google Scholar
Care AS, Bourque SL, Morton JS, Hjartarson EP, Robertson SA, Davidge ST. Снижение регуляторных Т-клеток на ранних сроках беременности вызывает дисфункцию матки у мышей.Гипертония. 2018; 72: 177–87.
CAS PubMed Google Scholar
Robertson SA, Care AS, Moldenhauer LM. Регуляторные Т-клетки при имплантации эмбриона и иммунный ответ на беременность. J Clin Invest. 2018; 128: 4225–35.
Google Scholar
Zhou CC, Zhang Y, Irani RA, Zhang H, Mi T, Popek EJ, et al. Аутоантитела, не зависящие от рецепторов ангиотензина, вызывают преэклампсию у беременных мышей.Nat Med. 2008; 14: 855–62 Epub 2008 27 июля.
CAS PubMed PubMed Central Google Scholar
Пэрриш М.Р., Мерфи С.П., Ратленд С., Уоллес К., Венцель К., Валлюкат Г. и др. Влияние иммунных факторов, фактора некроза опухоли альфа и агонистических аутоантител к рецептору ангиотензина II типа I на продукцию растворимого fms-подобного тирозина-1 и растворимого эндоглина в ответ на гипертензию во время беременности. Am J Hypertens.2010; 23: 911–6 Epub 2010, 29 апреля.
CAS PubMed PubMed Central Google Scholar
Вальтер Т., Янк А., Герингер-Вальтер С., Хорн Л.С., Степан Х. Рецептор ангиотензина II типа 1 оказывает влияние на плаценту мышей. Плацента. 2008; 29: 905–9.
CAS PubMed Google Scholar
Geusens N, Hering L, Verlohren S, Luyten C, Drijkoningen K, Taube M, et al.Изменения в эндоваскулярной инвазии трофобластов и ремоделировании спиральной артерии в срок в модели трансгенных преэкламптических крыс. Плацента. 2010. 31: 320–6.
CAS PubMed Google Scholar
Quitterer U, Fu X, Pohl A, Bayoumy KM, Langer A, Abdalla S. Бета-аррестин 1 предотвращает преэклампсию за счет подавления гетеромеров механочувствительных рецепторов AT1-B2. Клетка. 2019; 176: 318–333.e19.
CAS PubMed Google Scholar
Альбрехт Э.Д., Пепе ГДж. Эндокринология беременности. В: Бранс Ю.В., Кюль Т.Дж., редакторы. Приматы, не относящиеся к человеку, в перинатальных исследованиях. Нью-Йорк: Джон Уайли и сыновья; 1988. с. 13–78.
Google Scholar
Malassine A, Frendo JL, Evain-Brion D. Сравнение развития плаценты и эндокринных функций на модели человека и мыши. Обновление Hum Reprod. 2003; 9: 531–9.
CAS PubMed Google Scholar
Osol G, Mandala M. Ремоделирование сосудов матки матери во время беременности. Физиология (Bethesda). 2009; 24: 58–71.
Google Scholar
McCarthy FP, Kingdom JC, Kenny LC, Walsh SK. Модели преэклампсии на животных; использование и ограничения. Плацента. 2011; 32: 413–9.
CAS PubMed Google Scholar
Georgiades P, Ferguson-Smith AC, Burton GJ.Сравнительная анатомия развития дефинитивной плаценты мыши и человека. Плацента. 2002; 23: 3–19.
CAS PubMed Google Scholar
Силва Дж. Ф., Серакидес Р. Внутриутробная миграция трофобластов: сравнительный взгляд на людей и грызунов. Cell Adhes Migr. 2016; 10: 88–110.
CAS Google Scholar
Маршалл С.А., Ханнан, штат Нью-Джерси, Елинич М., Нгуен Т.П.Х, Гирлинг Дж.Э., Парри Л.Дж.Модели преэклампсии на животных: трансляционные ошибки и почему. Am J Physiol Regul Integr Comp Physiol. 2018; 314: R499–508.
CAS PubMed Google Scholar
Adamson SL, Lu Y, Whiteley KJ, Homyard D, Hemberger M, Pfarrer C, et al. Взаимодействие между клетками трофобласта и кровообращением матери и плода в плаценте мыши. Dev Biol. 2002; 250 (2): 358–73.
CAS PubMed Google Scholar
Schlabritz-Loutsevitch NE, Hubbard GB, Jenkins SL, Martin HC, Snider CS, Frost PA, et al. Онтогенез гематологических клеток и биохимические профили у бабуинов матери и плода (виды Papio). J Med Primatol. 2005; 34: 193–200.
CAS PubMed Google Scholar
Enders AC, Blankenship TN, Fazleabas AT, Jones CJ. Структура закрепляющих ворсинок и трофобластической оболочки плаценты человека, павиана и макаки.Плацента. 2001; 22: 284–303.
CAS PubMed Google Scholar
VandeBerg JL, Williams-Blangero S. Преимущества и недостатки нечеловеческих приматов в качестве животных моделей в генетических исследованиях сложных заболеваний. J Med Primatol. 1997; 26: 113–9.
CAS PubMed Google Scholar
Cox LA, Mahaney MC, Vandeberg JL, Rogers J. Карта генетического сцепления второго поколения генома павиана (Papio hamadryas).Геномика. 2006; 88: 274–81.
CAS PubMed Google Scholar
Кавана Д., Рао П.С., Цай С.К., О’Коннор ТК. Экспериментальная токсемия у беременных приматов. Am J Obstet Gynecol. 1977; 128: 75–85.
CAS PubMed Google Scholar
Макрис А., Торнтон С., Томпсон Дж., Томсон С., Мартин Р., Огл Р. и др. Ишемия матки и плаценты приводит к протеинурической гипертензии и повышению sFLT-1.Kidney Int. 2007. 71: 977–84.
CAS PubMed Google Scholar
Макрис А., Йунг К.Р., Лим С.М., Сандерленд Н., Хеффернан С., Томпсон Дж. Ф. и др. Фактор роста плаценты снижает кровяное давление на модели преэклампсии с маточно-плацентарной ишемией у нечеловеческих приматов. Гипертония. 2016; 67: 1263–72.
CAS PubMed PubMed Central Google Scholar
Туранов А.А., Ло А., Хасслер М.Р., Макрис А., Ашар-Патель А., Альтерман Дж. Ф. и др. Модуляция РНКи плацентарного sFLT1 для лечения преэклампсии. Nat Biotechnol. 2018; 36: 1164–1173.
Бонагура Т.В., Пепе Г.Дж., Эндерс А.С., Альбрехт Э.Д. Подавление экспрессии фактора роста эндотелия сосудов вневорсинчатого трофобласта и инвазии спиральной артерии матки эстрогеном во время ранней беременности павиана. Эндокринология. 2008. 149: 5078–87.
CAS PubMed PubMed Central Google Scholar
Бонагура Т.В., Бабищкин Я.С., Абердин Г.В., Пепе Г.Дж., Альбрехт Э.Д. Преждевременное повышение уровня эстрадиола на ранних сроках беременности бабуина подавляет ремоделирование маточных артерий и экспрессию вневорсинчатого фактора роста эндотелия сосудов плаценты и интегринов α1β1 и α5β1. Эндокринология. 2012; 153: 2897–906.
CAS PubMed PubMed Central Google Scholar
Aberdeen GW, Bonagura TW, Harman CR, Pepe GJ, Albrecht ED.Подавление ремоделирования маточной спиральной артерии трофобласта эстрогеном при беременности бабуином: влияние на динамику кровотока матки и плода. Am J Physiol Heart Circ Physiol. 2012; 302: h2936–44.
CAS PubMed PubMed Central Google Scholar
Babischkin JS, Aberdeen GW, Lindner JR, Bonagura TW, Pepe GJ, Albrecht ED. Доставка фактора роста эндотелия сосудов к базальной пластине плаценты способствует ремоделированию маточной артерии у приматов.Эндокринология. 2019; 160: 1492–505.
CAS PubMed PubMed Central Google Scholar
Аплин Дж. Д., Хей Т., Джонс С. Дж., Черч Х. Дж., Виовак Л. Развитие колонок цитрофобластов из эксплантированных ворсин плаценты человека в первом триместре: роль фибронектина и интегрина α5β1. Биол Репрод. 1999; 60: 828838.
Google Scholar
Ирвинг Дж. А., Лала П. К.. Функциональная роль интегринов клеточной поверхности в миграции клеток трофобласта человека: регуляция с помощью TGFβ, IGF-II и IGFBP-1.Exp Cell Res. 1995; 217: 419427.
Google Scholar
Фукусима К., Миямото С., Цукимори К., Кобаяси Х., Секи Х., Такеда С. и др. Фактор некроза опухоли и фактор роста эндотелия сосудов индуцируют репертории эндотелиального интегрина, регулируя эндоваскулярную дифференцировку и апоптоз в клеточной линии вневорсинчатого трофобласта человека. Биол Репрод. 2005; 73: 172–9.
CAS PubMed Google Scholar
Роль корина в инвазии трофобластов и ремоделировании спиральной артерии матки при беременности
Pijnenborg, R., Vercruysse, L. & Hanssens, M. Спиральные артерии матки при беременности человека: факты и противоречия. Плацента 27 , 939–958 (2006)
CAS Статья Google Scholar
Red-Horse, K. et al. Дифференциация трофобластов во время имплантации эмбриона и формирование границы раздела матери и плода. J. Clin. Вкладывать деньги. 114 , 744–754 (2004)
CAS Статья Google Scholar
Wu, Q., Xu-Cai, Y.O., Chen, S. & Wang, W. Corin: новое понимание натрийуретической пептидной системы. Kidney Int. 75 , 142–146 (2009)
CAS Статья Google Scholar
Yan, W., Sheng, N., Seto, M., Morser, J. & Wu, Q. Corin, мозаичная трансмембранная сериновая протеаза, кодируемая новой кДНК из сердца человека. J. Biol. Chem. 274 , 14926–14935 (1999)
CAS Статья Google Scholar
Лайн, К.Ю. и Робертс, Дж. М. Современные концепции патогенеза и лечения преэклампсии. J. Am. Med. Доц. 287 , 3183–3186 (2002)
Артикул Google Scholar
Сибай, Б., Деккер, Г. и Купферминц, М. Преэклампсия. Ланцет 365 , 785–799 (2005)
Артикул Google Scholar
Brosens, I.А., Робертсон, В. Б. и Диксон, Х. Г. Роль спиральных артерий в патогенезе преэклампсии. Акушерство. Гинеколь. Анну. 1 , 177–191 (1972)
CAS PubMed Google Scholar
Кауфманн, П., Блэк, С. и Хуппертц, Б. Эндоваскулярная инвазия трофобластов: последствия для патогенеза задержки внутриутробного развития и преэклампсии. Biol. Репродукция. 69 , 1–7 (2003)
CAS Статья Google Scholar
Norwitz, E.Р., Шуст, Д. Дж. И Фишер, С. Дж. Имплантация и выживание на ранних сроках беременности. N. Engl. J. Med. 345 , 1400–1408 (2001)
CAS Статья Google Scholar
Kanasaki, K. et al. Дефицит катехол-О-метилтрансферазы и 2-метоксиэстрадиола связан с преэклампсией. Природа 453 , 1117–1121 (2008)
ADS CAS Статья Google Scholar
Левин, Р.J. et al. Циркулирующие ангиогенные факторы и риск преэклампсии. N. Engl. J. Med. 350 , 672–683 (2004)
CAS Статья Google Scholar
Redman, C. W. & Sargent, I. L. Последние достижения в понимании преэклампсии. Наука 308 , 1592–1594 (2005)
ADS CAS Статья Google Scholar
Венкатеша, С.и другие. Растворимый эндоглин вносит свой вклад в патогенез преэклампсии. Nature Med. 12 , 642–649 (2006)
CAS Статья Google Scholar
Zhou, C.C. et al. Агонистические аутоантитела к рецепторам ангиотензина вызывают преэклампсию у беременных мышей. Nature Med. 14 , 855–862 (2008)
CAS Статья Google Scholar
Янь В., Wu, F., Morser, J. & Wu, Q. Corin, трансмембранная сердечная сериновая протеаза, действует как про-предсердный натрийуретический пептид-превращающий фермент. Proc. Natl Acad. Sci. США 97 , 8525–8529 (2000)
ADS CAS Статья Google Scholar
Chan, J. C. et al. Гипертензия у мышей, лишенных предсердной натрийуретической пептидной конвертазы корин. Proc. Natl Acad. Sci. США 102 , 785–790 (2005)
ADS CAS Статья Google Scholar
Дрис, Д.L. et al. Минорный аллель гена Корина, определяемый двумя миссенс-мутациями, часто встречается у чернокожих и связан с высоким кровяным давлением и гипертонией. Тираж 112 , 2403–2410 (2005)
CAS Статья Google Scholar
Davisson, R. L. et al. Открытие спонтанной генетической модели преэклампсии на мышах. Гипертония 39 , 337–342 (2002)
CAS Статья Google Scholar
Джон С.W. et al. Генетическое снижение предсердного натрийуретического пептида и солеочувствительная гипертензия. Наука 267 , 679–681 (1995)
ADS CAS Статья Google Scholar
Kuhn, M. et al. Натрийуретический пептид / гуанилциклаза — система функционирует как чувствительный к стрессу регулятор ангиогенеза у мышей. J. Clin. Вкладывать деньги. 119 , 2019–2030 (2009)
CAS Статья Google Scholar
Токудоме, Т.и другие. Нарушение восстановления кровотока после ишемии задних конечностей у мышей, лишенных гуанилилциклазы-А, рецептора натрийуретических пептидов предсердий и головного мозга. Артериосклер. Тромб. Васк. Биол. 29 , 1516–1521 (2009)
CAS Статья Google Scholar
Донг Н. и др. Плазморастворимый корин у пациентов с сердечной недостаточностью. Circ. Сердечная недостаточность. 3 , 207–211 (2010)
Статья Google Scholar
Цзян, Дж.и другие. Шеддинг эктодоменов и аутодективация протеазы корин сердечной мембраны. J. Biol. Chem. 286 , 10066–10072 (2011)
CAS Статья Google Scholar
Pan, J. et al. Геномные структуры генов корина человека и мыши и функциональные элементы GATA в их промоторах. J. Biol. Chem. 277 , 38390–38398 (2002)
CAS Статья Google Scholar
Knappe, S., Wu, F., Madlansacay, M. R. & Wu, Q. Идентификация доменных структур в пропептиде корина, необходимого для процессинга проатриального натрийуретического пептида. J. Biol. Chem. 279 , 34464–34471 (2004)
CAS Статья Google Scholar
Wang, W. et al. Вариант Корина, связанный с гипертонией и гипертрофией сердца, проявляет нарушенную активацию зимогена и активность процессинга натрийуретического пептида. Circ. Res. 103 , 502–508 (2008)
CAS Статья Google Scholar
Айронс, Д. У., Бейлис, П. Х., Батлер, Т. Дж. И Дэвисон, Дж. М. Предсердный натрийуретический пептид при преэклампсии: метаболический клиренс, экскреция натрия и почечная гемодинамика. Am. J. Physiol. 273 , F483 – F487 (1997)
CAS PubMed Google Scholar
Тихтонен, К.M., Koobi, T., Vuolteenaho, O., Huhtala, H. S. & Uotila, J. T. Натрийуретические пептиды и гемодинамика при преэклампсии. Am. J. Obstet. Гинеколь. 196 , 328.e1–328.e7 (2007)
Артикул Google Scholar
Chen, S. et al. Экспрессия и активность протеазы корина при сердечной недостаточности. Am. J. Physiol. Heart Circ. Physiol. 299 , h2687 – h2692 (2010)
CAS Статья Google Scholar
Ляо, Х., Wang, W., Chen, S. & Wu, Q. Роль гликозилирования в активации зимогена корина. J. Biol. Chem. 282 , 27728–27735 (2007)
CAS Статья Google Scholar
Донг Н. и др. Влияние антикоагулянтов на уровни растворимого корина в плазме крови человека, измеренные с помощью ELISA. Clin. Чим. Acta 411 , 1998–2003 (2010)
CAS Статья Google Scholar
Ву, Ф., Yan, W., Pan, J., Morser, J. & Wu, Q. Обработка про-предсердного натрийуретического пептида корином в сердечных миоцитах. J. Biol. Chem. 277 , 16900–16905 (2002)
CAS Статья Google Scholar
Qi, X., Jiang, J., Zhu, M. & Wu, Q. Изоформы корина человека с разными цитоплазматическими хвостами, которые изменяют нацеливание на клеточную поверхность. J. Biol. Chem. 286 , 20963–20969 (2011)
CAS Статья Google Scholar
Крюппель-подобный фактор 17 стимулирует экспрессию корки матки и способствует ремоделированию спиральной артерии во время беременности
Значение
Во время беременности кровеносные сосуды в матке увеличиваются в размере, обеспечивая больший приток крови и питательных веществ для растущего плода.Корин — это фермент, который ускоряет этот процесс. В этом исследовании мы стремились понять, как контролируется выработка корина матки. В экспериментах с человеческими маточными клетками и беременными мышами мы идентифицировали белок, названный Krüppel-подобным фактором 17 (KLF17), который включает ген корина в беременной матке. У генетически модифицированных мышей дефицит Klf17 предотвращает выработку корина матки, уменьшает размер маточных сосудов и вызывает высокое кровяное давление во время беременности. Эти результаты показывают, что KLF17 является ген-контролирующим белком, важным для функции матки во время беременности.
Abstract
Ремоделирование спиральной артерии — важный физиологический процесс беременной матки, который увеличивает приток крови к плоду. Нарушение ремоделирования спиральной артерии способствует преэклампсии, серьезному заболеванию беременных. Корин, трансмембранная сериновая протеаза, активируется в беременной матке, что способствует ремоделированию спиральной артерии. На сегодняшний день механизм, лежащий в основе активации корки матки, остается неизвестным. Здесь мы показываем, что Krüppel-подобный фактор (KLF) 17 является ключевым фактором транскрипции для экспрессии корина матки во время беременности.В культивируемых клетках эндометрия матки человека KLF17 связывается с промотором CORIN и усиливает активность промотора. Нарушение гена KLF17 в клетках эндометрия отменяет экспрессию CORIN . У мышей Klf17 активируется в беременной матке. Дефицит Klf17 предотвращает экспрессию в матке Corin во время беременности. Более того, мыши с дефицитом Klf17 и имеют плохо реконструированные спиральные артерии матки и развивают гестационную гипертензию и протеинурию.Вместе наши результаты показывают важную функцию KLF17 в регулировании экспрессии Corin и физиологии матки во время беременности.
Во время беременности спиральные артерии матки подвергаются процессу ремоделирования, в ходе которого гладкомышечные клетки в стенке сосуда и эндотелиальные клетки, выстилающие просвет, заменяются трофобластами, полученными из плаценты (1). Ремоделированные спиральные артерии сильно расширены, что снижает сопротивление сосудов матери и увеличивает кровоснабжение плода.Преэклампсия — это заболевание, которым страдают от 5 до 8% беременных женщин и вызывают смерть матери и плода (2). Нарушение ремоделирования спиральной артерии является основным фактором патогенеза преэклампсии (2-4). На сегодняшний день молекулярные механизмы, лежащие в основе ремоделирования спиральной артерии, полностью не изучены.
Предсердный натрийуретический пептид (ПНП) - это сердечный гормон, регулирующий водно-солевой баланс и артериальное давление (5). Он синтезируется в виде предшественника, то есть про-ANP, который превращается в зрелый ANP с помощью корина, трансмембранной сериновой протеазы, высоко экспрессируемой в сердце (6, 7).Помимо сердца, корин и ПНП экспрессируются в тканях вне сердца, таких как почки (8⇓ – 10), волосяные фолликулы (11, 12) и развивающиеся кости (13, 14). Недавно мы и другие показали, что корин активируется в беременной матке, где корин и ANP способствуют инвазии трофобластов и ремоделированию спиральной артерии (15, 16). У беременных мышей дефицит корина и ANP нарушает ремоделирование спиральной артерии, вызывая гестационную гипертензию и протеинурию, фенотип, подобный преэклампсии (15, 17, 18).У людей, у женщин с преэклампсией сообщалось о снижении уровней информационной РНК (мРНК) и белка корина, а также о вредоносных вариантах CORIN (15, 19, 20). Эти данные показывают, что повышение регуляции корина матки является частью физиологической реакции при нормальной беременности. Однако остается неизвестным, как регулируется экспрессия корина в беременной матке.
В сердце экспрессия натрийуретического пептида контролируется GATA-4, фактором транскрипции цинкового пальца (21, 22). Сходный GATA-4-зависимый механизм является критическим для экспрессии сердечного корина.Мы показали, что промоторы генов корина человека и мыши содержат консервативную последовательность, которая необходима для связывания GATA-4 и экспрессии корина в кардиомиоцитах (23). GATA-4 в первую очередь является фактором транскрипции сердца, но не экспрессируется в матке (24). Вероятно, что транскрипционный механизм, контролирующий экспрессию корина в матке, отличается от такового в сердце.
Чтобы проверить эту гипотезу, мы проанализировали активность промотора CORIN в клетках эндометрия и сердца.Мы также исследовали экспрессию корина и фактора транскрипции в децидуализированных клетках эндометрия человека и беременных матках мышей. Мы определили Krüppel-подобный фактор 17 (KLF17) как ключевой фактор транскрипции для экспрессии корина матки. У мышей дефицит Klf17 предотвращал экспрессию Corin в беременной матке. У Klf17 -дефицитных мышей развилась индуцированная беременностью гипертензия и протеинурия. Эти находки указывают на важную роль KLF17 в экспрессии Corin и физиологии матки во время беременности.
Результаты
Человек
Активность промотора CORIN в маточных и сердечных клетках.Ранее мы идентифицировали консервативную GATA-связывающую последовательность в промоторе CORIN человека, необходимую для экспрессии корина в кардиомиоцитах (23). В этом исследовании мы проанализировали фрагмент промотора CORIN длиной 405 п.н., содержащий GATA-связывающую последовательность (рис. 1 A ). В анализе люциферазы промоторная активность была обнаружена в клетках AN3-CA эндометрия матки человека и кардиомиоцитах HL-1 мыши, в которых сообщалось об экспрессии эндогенного корина (25, 26) (рис.1 B и C ). Внутри этого промоторного фрагмента сайт связывания E-box и два сайта связывания KLF располагались выше сайта связывания GATA (рис. 1 A и SI, приложение , рис. S1). Делеция сайта связывания E-box оказывала незначительное влияние на промоторную активность CORIN в клетках AN3-CA и HL-1 (рис. 1 A — C ). Дальнейшая делеция двух сайтов связывания KLF устраняет активность промотора в клетках AN3-CA, но не в HL-1 (рис.1 A — C ), предполагая возможную роль сайтов связывания KLF в экспрессии корина в клетках эндометрия матки.
Рис. 1.Активность промотора CORIN в клетках эндометрия и кардиомиоцитах матки. ( A ) Иллюстрация конструкции люциферазы с фрагментом промотора CORIN длиной 405 п.н. с сайтом связывания E-box, двумя сайтами связывания KLF и сайтом связывания GATA (P405) и двумя конструкциями с 236- и 161 п.н., соответственно, усеченные фрагменты промотора CORIN (P236 и P161).( B и C ) Люциферазная активность в клетках эндометрия AN3-CA ( B ) и HL-1 ( C ), трансфицированных конструкциями промотора CORIN . Конструкцию pGL3 использовали в качестве фонового контроля. ( D ) Иллюстрация промоторных конструкций CORIN с мутациями (жирным курсивом) в одном (KM1 и KM2) или обоих (KM3) сайтах связывания KLF. ( E и F ) Люциферазная активность в клетках AN3-CA ( E ) и HL-1 ( F ), трансфицированных конструкциями промотора CORIN .( G ) Иллюстрация конструкции промотора CORIN с мутациями (жирные курсивные буквы) в сайте связывания GATA (GM). ( H и I ) Люциферазная активность в клетках AN3-CA ( H ) и HL-1 ( I ), трансфицированных конструкциями промотора CORIN . n = от 3 до 6 на группу. Данные представлены как среднее ± стандартная ошибка среднего; Значения P анализировали с помощью двустороннего критерия Стьюдента t . нс, не имеет значения.
Чтобы проверить наши результаты, мы мутировали сайты связывания KLF, по отдельности или вместе (рис. 1 D ). В анализе люциферазы промоторный фрагмент с мутациями в отдельных сайтах связывания KLF имел пониженную активность в клетках AN3-CA (фиг. 1 E ). Когда оба сайта связывания KLF были мутированы, активность промотора отменялась в клетках AN3-CA, но не в HL-1 (фиг. 1 E и F ). Напротив, мутации в сайте связывания GATA отменяли промоторную активность в клетках HL-1, но не в клетках AN3-CA (рис.1 G — I ). Эти результаты показывают, что экспрессия корина в клетках эндометрия матки контролируется транскрипционным механизмом с участием фактора (ов) транскрипции KLF, но не GATA.
Экспрессия корина матки и KLF у мышей, получавших прогестерон, и беременных мышей.
Семья KLF состоит из 17 членов (27). Чтобы идентифицировать члена (ов) KLF, ответственного за экспрессию корина в беременной матке, мы обработали овариэктомированных мышей C57BL / 6 эстрогеном или прогестероном ( SI Приложение , рис.S2 A ). С помощью qRT-PCR мы обнаружили повышенные уровни мРНК Corin в матке у мышей, получавших прогестерон, но не получавших эстроген, по сравнению с таковыми у мышей, получавших носитель (фиг. 2 A ). Затем мы проанализировали экспрессию в матке всех 17 генов Klf с помощью qRT-PCR у овариэктомированных мышей, получавших прогестерон. Мы обнаружили повышенные уровни мРНК в матке Corin , Klf2 , Klf9 , Klf15 и Klf17 , но не остальных генов Klf (рис.2 B и SI Приложение , рис. S2 B ). Мы подтвердили эти результаты с помощью анализа qRT-PCR тканей матки беременных мышей C57BL / 6 на разных (G) днях гестации (рис. 2 C ). Эти результаты показывают, что Klf2, Klf9, Klf15 и Klf17 являются возможными кандидатами, ответственными за активацию корина в беременной матке.
Рис. 2. ЭкспрессияCorin и Klf в матках мышей. ( A ) Уровни мРНК Corin в матке от овариэктомированных мышей, получавших носитель, эстроген или прогестерон.( B ) Уровни мРНК Corin , Klf2 , Klf9 , Klf15 и Klf17 в матках от овариэктомированных мышей, получавших носитель или прогестерон. ( C ) Уровни мРНК Corin , Klf2 , Klf9 , Klf15 и Klf17 в матках небеременных (NP) и беременных мышей C57BL / 6 на 4,5, 7,5 и 12,5 гестационных днях. n = от 4 до 5 на группу в A ; n = 10 на группу в B ; n = от 9 до 10 на группу в C .Данные представлены как среднее ± стандартная ошибка среднего; Значения P анализировали с помощью ANOVA ( A, и C ) или двустороннего теста Стьюдента t ( B ). нс, не имеет значения.
Влияние экспрессии KLF на активность промотора
CORIN в клетках матки.Для изучения функции кандидатов в KLF мы котрансфицировали клетки AN3-CA с промоторными конструкциями CORIN и плазмидами, экспрессирующими человеческие белки KLF2, KLF9, KLF15 и KLF17.Повышенная промоторная активность была обнаружена, когда плазмиды, экспрессирующие KLF2 и KLF17, но не KLF9 и KLF15, котрансфицировали промоторной конструкцией с сайтами связывания KLF (фиг. 3 A и B ). Путем вестерн-блоттинга мы подтвердили экспрессию рекомбинантного KLF в трансфицированных клетках (фиг. 3 C ). В этих экспериментах промоторная активность не увеличивалась, если котрансфекция проводилась с использованием промоторных конструкций CORIN с удаленными сайтами связывания KLF (рис.3 D ) или мутировавший (рис. 3 E ). В отличие от клеток AN3-CA, экспрессия рекомбинантных KLF2 и KLF17 в кардиомиоцитах HL-1 не увеличивала активность промотора при использовании конструкций CORIN с сайтами связывания KLF или без них ( SI Приложение , рис. S3 A — D ). Эти результаты показывают, что KLF2 и / или KLF17, вероятно, ответственны за активацию корина в беременной матке.
Рис. 3.Влияние экспрессии рекомбинантного белка KLF на активность промотора CORIN в клетках AN3-CA эндометрия матки.( A ) Иллюстрация использованных промоторных конструкций CORIN . ( B ) Люциферазная активность в клетках AN3-CA, котрансфицированных конструкцией промотора P236 CORIN и контрольным вектором (Vec) или плазмидами, экспрессирующими человеческий KLF2 (K2), KLF9 (K9), KLF15 (K15) или KLF17 ( К17). n = 8 на группу. Данные представлены как среднее ± стандартная ошибка среднего; P Значения анализировали с помощью ANOVA. нс, не имеет значения. ( C ) Вестерн-блоттинг рекомбинантных белков KLF в трансфицированных клетках с использованием антитела против FLAG.( D и E ) Люциферазная активность в клетках AN3-CA, трансфицированных конструкцией промотора P405 CORIN (положительный контроль) или котрансфицированных промотором P161 ( D ) или KM3 ( E ) промотор CORIN конструкция и контрольный вектор (Vec) или плазмиды, экспрессирующие человеческий KLF2 (K2), KLF9 (K9), KLF15 (K15) или KLF17 (K17). n = 3 на группу. Данные представлены как среднее ± стандартная ошибка среднего; P Значения (по сравнению с P405) и нс (несущественно) (vs.группа, котрансфицированная вектором) были проанализированы с помощью ANOVA.
Повышенная экспрессия корина и KLF17 в децидуализированных стромальных клетках эндометрия человека.
Стромальные клетки эндометрия человека (HESC) обычно используются для индукции децидуализации в культуре, чтобы имитировать клетки эндометрия в беременной матке (28). Мы культивировали HESC в среде для децидуализации и обнаружили повышенные уровни мРНК PRL (кодирующий пролактин, маркер децидуализации) и CORIN с помощью qRT-PCR (рис.4 A ), что соответствует повышенной регуляции корина в матке беременной (15). Посредством вестерн-блоттинга были обнаружены повышенные уровни KLF17 в децидуализированных HESC (фиг. 4 B ). Напротив, уровни KLF2 не изменились в обработанных аналогичным образом HESC (фиг. 4 C ). Мы подтвердили эти результаты с помощью иммуногистохимии (фиг. 4 D ) и коиммунофлуоресцентного окрашивания (фиг. 4 E ), которые показали повышенное окрашивание цитоплазмы и мембран корина и ядерное окрашивание KLF17 в децидуализированных, но не контрольных, HESC.В соответствии с результатами вестерн-блоттинга иммуногистохимия показала сходные уровни ядерного окрашивания KLF2 в HESC до и после децидуализации ( SI, приложение , рис. S4).
Рис. 4. ЭкспрессияKLF17 и KLF2 в децидуализированных HESC. ( A ) Культивированные HESC индуцировали децидуализацию. Уровни мРНК PRL ( верхний, ) и CORIN ( нижний ) в указанные моменты времени анализировали с помощью qRT-PCR. ( B и C ) Вестерн-блоттинг эндогенных белков KLF17 ( B ) и KLF2 ( C ) в HESC, культивируемых в децидуализационной среде, с течением времени.GAPDH использовали в вестерн-блоттинге ( верх, ) для расчета относительных уровней экспрессии белка ( низ ). n = 5 на группу в A ; n = 3 на группу в B и C . На гистограммах данные представлены как среднее ± SEM; нс, не имеет значения; ** P <0,01 по сравнению с контролем в момент времени 0 в том же эксперименте, согласно анализу ANOVA. ( D и E ) Иммуногистохимия ( D ) и коиммунофлуоресцентное окрашивание ( E ) эндогенных белков корина и KLF17 в контрольных (неиндуцированных) и децидуализированных (индуцированных) HESC.В D нормальный IgG использовали в качестве отрицательного контроля. Данные представляют из трех независимых экспериментов. (Шкала 20 мкм.)
Иммунопреципитационный анализ хроматина.
Чтобы проверить, связывается ли KLF17 непосредственно с промотором CORIN , мы выполнили анализ иммунопреципитации хроматина (ChIP) в HESC до и после децидуализации. Положительные фрагменты ПЦР были обнаружены, когда ChIP проводился с антителом против KLF17 в децидуализированных, но не контрольных HESC (рис.5 A и B ). Напротив, при использовании контрольного IgG или антитела против KLF2 или против KLF9 не было обнаружено никаких ПЦР-фрагментов (фиг. 5 B ). В положительном контроле фрагменты ПЦР были обнаружены, когда ChIP проводился с антителом против гистона 3 в HESC до и после децидуализации (фиг. 5 B ). Результаты были подтверждены дополнительным анализом qPCR ( SI, приложение , рис. S5, A ), показывающим, что KLF17, но не KLF2, связывается непосредственно с промотором CORIN в децидуализированных HESC.
Рис. 5. АнализChIP в HESC и экспрессия CORIN в HESC KLF17 -KO. ( A ) Иллюстрация анализа ChIP, в котором фрагменты хроматина из неиндуцированных и индуцированных HESC осаждали антителами против гистона 3 (h4) (положительный контроль) или белков KLF и использовали для ПЦР с праймерами для сайтов, отличных от KLF и KLF. ( B ) Продукты ПЦР представлены в агарозных гелях. ( C ) Ген KLF17 был разрушен в HESC с использованием CRISPR / Cas9.Показаны данные частичного секвенирования HESC WT и KLF17 -KO. Красный прямоугольник указывает на удаленную последовательность. ( D ) Вестерн-блоттинг эндогенного белка KLF17 в HESC WT и KLF17 -KO без (-) или с (+) децидуализацией (индукцией). GAPDH был контролем. Представлены несмежные дорожки из того же вестерн-блоттинга. Данные представляют из трех независимых экспериментов. ( E ) Относительные уровни мРНК CORIN в HESC WT и KLF17 -KO без (неиндуцированной) или с (индуцированной) децидуализацией. n = 5 на группу. Данные представлены как среднее ± стандартная ошибка среднего; Значения P анализировали с помощью двустороннего критерия Стьюдента t . нс, не имеет значения.
Corin Expression in
KLF17 -Knockout HESCs.Чтобы определить функцию KLF17 в повышающей регуляции экспрессии корина в клетках эндометрия матки, мы разрушили ген KLF17 в HESC с помощью CRISPR / Cas9 (рис. 5 C ). При вестерн-блоттинге белок KLF17 не был обнаружен в HESC KLF17 -KO до или после децидуализации (рис.5 D ). В qRT-PCR в децидуализированных HESC были обнаружены сильно повышенные уровни мРНК CORIN по сравнению с таковыми в контрольных клетках (фиг. 5 E ). Такая повышающая регуляция, однако, не была обнаружена в HESC KLF17 -knockout (KO) (рис. 5 E ). Мы подтвердили эти результаты в отдельной линии HESC KLF17 -KO, в которой ген KLF17 был разрушен в другом сайте экзона 1 ( SI Приложение , рис. S5 B — D ).Эти данные указывают на критическую роль KLF17 в активации CORIN в децидуализированных клетках эндометрия матки человека.
Поколение
Klf17 — / — Мышей.Чтобы понять функциональную важность Klf17 in vivo, мы использовали эмбриональные стволовые клетки мыши с нарушенным аллелем Klf17 для получения мышей Klf17 — / — ( SI Приложение , рис. S6 A ). и B ). Klf17 — / — мыши были жизнеспособны без заметных физических отклонений.При вскрытии трупов и гистологических исследованиях не было обнаружено явных дефектов в основных органах, включая семенник, в котором обнаружена экспрессия Klf17 (29). При ОТ-ПЦР повышающая регуляция Klf17 и Corin в матке была обнаружена у беременных мышей дикого типа (WT), но не Klf17 — / — , мышей (фиг. 6, A ). Соответственно, qRT-PCR показала заметное снижение уровней мРНК Corin в матке у беременных мышей Klf17 — / — по сравнению с таковыми у беременных мышей WT (рис.6 В ). Результаты были подтверждены вестерн-блоттингом белков Klf17 и Corin в образцах матки мышей WT и Klf17 — / — ( SI Приложение , рис. S7, A, и B ). Напротив, сердечные уровни мРНК Corin были сходными у небеременных и беременных мышей WT и Klf17 — / — в анализах ОТ-ПЦР и qRT-PCR ( SI Приложение , Рис. S7, C и D ).
Рис. 6. Экспрессия генаи фертильность у мышей Klf17 — / — . ( A ) Уровни мРНК Klf17 и Corin в матках небеременных (NP) и беременных (G12.5 и G18.5) мышей WT и Klf17 — / — (KO) мышей, как анализировали с помощью ОТ-ПЦР. Образцы, в которых не использовались матрицы кДНК, были отрицательными контролями (Ctr). ( B ) Относительные уровни мРНК Corin у беременных (G12.5 и G18.5) Мыши WT и KO по результатам анализа с помощью qRT-PCR. n = от 8 до 14 на группу. ( C ) Количество и процентное соотношение Klf17 + / + , Klf17 +/− и Klf17 — / — щенков в 28 помётах из 18 Klf17 +/− мышей. ( D ) Количество детенышей в помете от вязок WT × WT ( n = 22) и KO × KO ( n = 52). ( E ) Репрезентативные изображения эмбрионов G12.5 самок WT и KO спаривались с самцами того же генотипа. Стрелки указывают на аномальные эмбрионы. ( F ) Репрезентативные изображения с большим увеличением (× 16), показывающие нормальный эмбрион от мыши WT и два эмбриона (один нормальный и один ненормальный) от мыши KO. Гистограмма показывает процент нормальных и аномальных эмбрионов в 90 эмбрионах от мышей KO. ( G ) Количество детенышей в помете от самок WT и KO, повязанных с самцами WT или KO. Данные представлены как среднее ± стандартная ошибка среднего; Значения P анализировали с помощью двустороннего теста Стьюдента t ( B и D ) или ANOVA ( G ).нс, не имеет значения.
Фертильность
Klf17 — / — самцов и самок мышей.Мы скрестили Klf17 +/- мышей и нашли ожидаемое менделевское соотношение Klf17 + / + , Klf17 +/- и Klf17 — / — потомков (рис. 6 C ), что указывает на отсутствие явной эмбриональной потери у беременных мышей Klf17 +/- .Спаривание Klf17 — / — самцов и самок мышей давали меньше детенышей на помет по сравнению с Klf17 + / + самцов и самок (4,1 ± 0,3 [ n = 52] по сравнению с 7,1 ± 0,4 [ n = 22]; P <0,01) (рис. 6 D ), что указывает на гибель плода у беременных мышей Klf17 — / — . Затем мы исследовали эмбрионы беременных мышей Klf17 + / + и Klf17 — / — , спарившихся с самцами того же генотипа.На G12,5 d общее количество эмбрионов на мышь у мышей Klf17 + / + и Klf17 — / — было одинаковым (7,2 ± 0,5 [ n = 11] против 7,6 ± 0,7 [ n = 17]; P = 0,64) (Рис. 6 E ), что предполагает аналогичное оплодотворение и имплантацию эмбриона между Klf17 + / + и Klf17 — / — мышей. Однако у мышей Klf17 — / — часть эмбрионов (29/90, 32.2%) были маленькими и аномальными, тогда как другие эмбрионы (61/90, 67,7%) были неотличимы по размеру и морфологии от таковых у мышей Klf17 + / + (Рис.6 E и F ). Гистологический анализ показал некроз тканей аномальных эмбрионов мышей Klf17 — / — ( SI Приложение , рис. S8). Из 128 обследованных эмбрионов Klf17 + / + мышей только 3 (3/128, 2.3%) оказались ненормальными.
Чтобы определить, была ли гибель плода вызвана дефицитом Klf17 у самцов или самок мышей, мы скрестили Klf17 + / + и Klf17 — / — мышей с мышами того же типа. или другой генотип. Щенков в помете от Klf17 — / — самок, вне зависимости от вязки с Klf17 + / + или Klf17 — / — самцов, было намного меньше, чем от Klf17 + / + самок, при вязке с Klf17 + / + или Klf17 — / — самцов (рис.6 G ). Эти результаты показывают, что гибель плода была вызвана дефицитом Klf17 у самок мышей, независимо от генотипа у спаривающихся самцов или полученных плодов, что указывает на важность Klf17 в матке беременной, но не в плаценте или других тканях плода.
Инвазия трофобластов и маточные спиральные артерии у мышей
Klf17 — / — .Корин способствует инвазии трофобластов и ремоделированию спиральной артерии в матке беременной (15). Мы провели гистологический и иммуногистохимический анализ у беременных мышей Klf17 + / + и Klf17 — / — .На G12,5 d мы обнаружили инвазию трофобластов, на что указывало окрашивание цитокератина, в децидуальной оболочке мышей Klf17 + / + (фиг. 7, A и B ). Для сравнения, инвазия трофобластов была заметно снижена в децидуальной оболочке мышей Klf17 — / — (фиг. 7, A, и B, ). На G18.5 d децидуальные артерии у мышей Klf17 + / + были больше в диаметре по сравнению с таковыми у мышей Klf17 — / — (рис.7 C и D ). В отличие от мышей Klf17 + / + , децидуальные артерии у мышей Klf17 — / — имели больше гладкомышечных клеток, на что указывало окрашивание α-гладкомышечного актина (SMA) (рис. 7 D ). Эти результаты показывают, что инвазия трофобластов и ремоделирование спиральной артерии матки нарушены у беременных мышей Klf17 — / — .
Рис. 7.Инвазия трофобластов и ремоделирование спиральной артерии у мышей Klf17 — / — .( A и B ) Плацентарные (P) и маточные (U) срезы мышей WT и Klf17 — / — (KO) на G12.5 окрашивали H&E ( A ) или иммунное окрашивание антителом против цитокератина 18 (Cytok) ( B ). Области в рамке на изображениях с малым увеличением (× 25) ( верхний ряд, ) показаны при большем увеличении (× 100) ( нижний ряд ). Эндоваскулярная инвазия трофобластов в децидуальных отделах указана стрелками.( C и D ) Срезы плаценты (P) и матки (U) мышей WT и KO на G18.5 были окрашены H&E ( C ) или иммуноокрашены антителами против SMA ( D ). Области в рамке изображений с низким увеличением (× 25) ( верхний ряд, ) показаны с большим увеличением (× 100) ( нижний ряд ). Положительное окрашивание SMA в децидуальных артериях у мышей KO указано стрелками. Данные являются репрезентативными для исследований, по крайней мере, на пяти мышах в каждой группе.
Гипертония и протеинурия, вызванные беременностью, у мышей
Klf17 — / — .Нарушение инвазии трофобластов и ремоделирование спиральной артерии способствуют преэклампсии, заболеванию, характеризующемуся гестационной гипертензией и протеинурией. Мы измерили артериальное давление у небеременных и беременных мышей Klf17 + / + и Klf17 — / — . До беременности или при G12,5 дн у мышей Klf17 + / + и Klf17 — / — артериальное давление было примерно одинаковым (рис.8 А ). На G18,5 дн артериальное давление у мышей Klf17 — / — увеличилось по сравнению с таковыми в небеременности или на G12,5 дн (121,2 ± 1,3 [ n = 12] по сравнению с 104,6 ± 1,5 [ n = 14] или 106,1 ± 2,3 [ n = 14] мм рт. 1,3 [ n = 12] по сравнению с 103,6 ± 2,8 [ n = 8] мм рт. Ст .; P <0.001) (рис.8 A ). Эти результаты показывают, что у мышей Klf17 — / — развивается гипертензия, вызванная беременностью.
Рис. 8.Гипертония, протеинурия и ишемические клубочки у мышей Klf17 — / — . ( A ) Систолическое артериальное давление (АД) у небеременных (NP) и беременных (G12.5 и G18.5) мышей WT и Klf17 — / — (KO). n = от 8 до 14 на группу. ( B ) Уровни белка в моче у NP и беременных (G12.5 и G18.5) мышей WT и KO. n = от 7 до 16 на группу. ( C ) Гистология клубочков на срезах почек, окрашенных PAS и трихромом по Массону, от мышей NP и беременных (G12.5 и G18.5) мышей WT и KO. (Шкала 20 мкм). ( D ) Гломерулярные эритроциты (RBC) подсчитывали в почечных срезах мышей NP и беременных (G12.5 и G18.5) мышей WT и KO. Для A , B и D данные представлены как среднее ± стандартная ошибка среднего; P Значения анализировали с помощью ANOVA; нс, не имеет значения.
Затем мы измерили уровни белка в моче у мышей Klf17 + / + и Klf17 — / — . Сходные уровни белка в моче были обнаружены у мышей Klf17 + / + и Klf17 — / — до беременности или на G12,5 d (рис. 8 B ). На G18,5 d уровень белка в моче увеличился у мышей Klf17 — / — , но не у Klf17 + / + , мышей (237.2 ± 12,9 [ n = 12] против 103,4 ± 21,2 [ n = 7] мг / дл; P <0,001) (Рис.8 B ). На 7 день послеродового периода уровень артериального давления и белка в моче у мышей Klf17 — / — снизился до аналогичных уровней у контрольных мышей Klf17 + / + ( SI Приложение , рис. S9) .
При гистологическом анализе морфология клубочков у небеременных и беременных Klf17 + / + мышей и небеременных Klf17 — / — мышей оказались нормальными при окрашивании периодической кислотой по Шиффу (PAS) и по Массону. Срезы почек, окрашенные трихромом (рис.8 С ). У беременных мышей Klf17 — / — , особенно на поздней стадии гестации (G18.5), наблюдались ишемические клубочки, о чем свидетельствуют капилляры меньшего диаметра и меньшее количество эритроцитов (рис. 8 C ). и D ). Эти данные показывают, что у мышей Klf17 — / — развивается ишемия почек во время беременности.
Обсуждение
В этом исследовании мы изучили механизм, лежащий в основе экспрессии корина в беременной матке.Мы определили KLF17 как ключевой фактор транскрипции для экспрессии корина матки во время беременности. Этот вывод подтверждается множеством экспериментальных данных из анализов промотора CORIN и ChIP, профилирования маточного белка Klf у овариэктомированных мышей, получавших прогестерон, и нормальных беременных мышей, нарушения гена KLF17 в HESC и характеристики беременных Klf17 — / — мышей. Наши результаты показывают KLF17-зависимый транскрипционный механизм экспрессии корина матки, который отличается от опосредованного GATA-4 механизма экспрессии сердечного корина (23).Протеазы, составляющие ~ 2% белков, кодируемых геномом человека (30), имеют большое биологическое значение. Корин представляет собой сериновую протеазу, подобную трипсину, которая, как было показано, обладает такими отчетливыми механизмами транскрипции в определенных тканях.
KLF17 является членом семейства KLF, группы структурно связанных факторов транскрипции, которые действуют в различных тканях, регулируя множество физиологических и патологических процессов (27, 31, 32). Как и все KLF, KLF17 содержит три консервативных C-концевых ДНК-связывающих цинковых пальца, которые распознают последовательности GC-box и CACCC-box (29, 33).По сравнению с другими KLF, KLF17 млекопитающих гораздо менее известен в отношении своей физиологической функции. Большинство исследований KLF17 сосредоточено на его роли в биологии опухолей (34, 35). Подавление KLF17 было идентифицировано при многих раковых заболеваниях человека, включая рак груди, легких, печени и желудка (34, 36–38). Было показано, что KLF17 подавляет эпителиально-мезенхимальный переход и метастазирование рака в трансформирующих сигнальных механизмах, опосредованных факторами роста β и p53 (34, 39, 40).
У рыбок данио и Xenopus ген klf17 экспрессируется в бластуле, невромастах боковой линии и выводных железах (41–43).У курицы эмбриональная экспрессия klf17 была обнаружена в первичной полоске (44). Нарушение гена klf17 у рыбок данио предотвращает вылупление эмбрионов, что приводит к летальному исходу (45). Сходные результаты были обнаружены у эмбрионов Xenopus , когда klf17 , также называемый Nepture , был заблокирован антисмысловыми олигонуклеотидами (46). Эти данные демонстрируют важность klf17 в эмбриогенезе ранних видов позвоночных. В отличие от рыб и Xenopus , у млекопитающих нет выводных желез.У мышей высокие уровни мРНК Klf17 (также называемой Zfp393 ) были обнаружены в сперматидах и развивающихся ооцитах (29). KLF17 Экспрессия мРНК также была обнаружена в семенниках человека (33), что указывает на потенциальную роль KLF17 млекопитающих в гаметогенезе. В нашем исследовании, однако, мы обнаружили, что и самцы, и самки мышей Klf17 — / — давали жизнеспособное потомство, что указывает на то, что Klf17 не нужен для сперматогенеза и ооцитогенеза у мышей.Более того, мыши Klf17 — / — и Klf17 + / + имели одинаковое количество эмбрионов на уровне G12.5, что указывает на то, что овуляция, оплодотворение и имплантация эмбриона не нарушены Klf17 . дефицит. Мы также наблюдали ожидаемое менделевское соотношение среди потомков от спаривания Klf17 +/- , что указывает на то, что Klf17 незаменим в эмбриогенезе.
члена KLF, включая KLF5, KLF9, KLF11 и KLF13, вовлечены в патофизиологию матки (47–52).У людей сверхэкспрессия KLF17 наблюдалась при поздних стадиях рака эндометрия (53). В этом исследовании мы обнаружили неожиданную роль KLF17 в матке беременной. Мы обнаружили, что при стимуляции прогестероном экспрессия KLF17 повышалась в клетках эндометрия матки человека и в матке беременных мышей, что было связано с повышенной экспрессией корина. Соответственно, беременные мыши Klf17 — / — лишены экспрессии корина матки. У мышей также была отсроченная инвазия трофобластов, плохо ремоделированные маточные спиральные артерии и меньший размер помета, аналогичный фенотип отмечен у мышей с дефицитом корина и ANP (15).Более того, у мышей Klf17 — / — развилась поздняя гестационная гипертензия и протеинурия, которые также наблюдались у мышей с дефицитом корина и ANP (15). Эти результаты подтверждают решающую роль KLF17 в экспрессии корина и биологии матки. Предполагается, что как фактор транскрипции KLF17 регулирует другие гены в беременной матке. Дальнейшие исследования важны для идентификации этих KLF17-зависимых генов и понимания их роли в регуляции физиологии матки во время беременности.
Таким образом, ремоделирование спиральной артерии матки является физиологическим механизмом беременности. Корин — это протеаза, которая способствует ремоделированию спиральной артерии беременной матки. Низкие уровни мРНК CORIN были идентифицированы у пациентов с преэклампсией (15), что указывает на потенциальную роль дефектной транскрипции CORIN в заболевании. Здесь мы показываем, что KLF17 необходим для активации CORIN в клетках эндометрия матки человека и что дефицит Klf17 устраняет экспрессию Corin в матке у мышей.Наши результаты показывают, что KLF17 является важным фактором транскрипции, который регулирует экспрессию корина и биологию матки во время беременности.
Материалы и методы
Плазмиды.
Плазмида светлячка на основе pGL3 (Promega), содержащая последовательность промотора CORIN человека длиной 405 пар оснований, и плазмида pRL-SV40, экспрессирующая люциферазу Renilla (Promega), были описаны ранее (23). Дополнительные плазмиды с последовательными делециями в промоторной последовательности CORIN получали с помощью сайт-направленного мутагенеза (Agilent Technologies).Плазмиды, экспрессирующие белки KLF2 (RC210042), KLF9 (RC210147), KLF15 (RC207523) и KLF17 (RC208293) человека, были получены от OriGene. Белки KLF, экспрессируемые плазмидами на основе pCMV6, содержали C-концевую метку Myc-DDK (FLAG) для обнаружения белка.
Культура клеток.
Мышиные кардиомиоциты HL-1, первоначально из Уильяма Клейкомба, Медицинский центр Университета штата Луизиана, Новый Орлеан, Лос-Анджелес (без контаминации микоплазмой), культивировали в среде Клейкомба (Sigma) с 10% фетальной бычьей сывороткой (FBS) и 4 мМ л. -глютамин.Клетки аденокарциномы эндометрия матки человека AN3-CA (ATCC HTB-111; профиль короткого тандемного повтора (STR); отсутствие контаминации микоплазмой) и HESC (ATCC CRL-4003; профиль STR; отсутствие контаминации микоплазмой) культивировали, соответственно, в минимальной среде Игла. (MEM) (HyClone, Sh40024.01B) с 10% FBS и средой Eagle, модифицированной Дульбекко (DMEM) / средой Ham’s F-12 (Sigma) с 10% FBS, очищенным от угля и декстрана (Biological Industries), 1,5 г / л бикарбоната натрия и 1% инсулина, трансферрина и селена (Sigma).Клетки культивировали при 37 ° C в увлажненных инкубаторах с 5% CO 2 .
Двойной репортерный анализ люциферазы.
Люциферазные плазмиды с фрагментами промотора CORIN человека и контрольную плазмиду pGL3 трансфицировали в клетки AN3-CA и HL-1 с помощью реагента Lipofectamine 2000 (Thermo Fisher). Плазмиду pRL-SV40, экспрессирующую люциферазу Renilla , котрансфицировали в качестве контроля эффективности трансфекции. Через 40 ч при 37 ° C трансфицированные клетки лизировали буфером, предоставленным в наборе Dual-Luciferase Reporter Assay (Promega).Через 15 мин при комнатной температуре добавляли буфер для анализа люциферазы и последовательно измеряли активность люциферазы светлячка и Renilla в ридере для микропланшетов Luminoskan Ascent (Thermo Fisher). Активность промотора CORIN рассчитывали на основании нормализованной активности люциферазы светлячков.
Вестерн-блоттинг рекомбинантных KLF в трансфицированных клетках.
Для исследования влияния экспрессии KLF на активность промотора CORIN плазмиды, экспрессирующие человеческие KLF2, KLF9, KLF15 и KLF17, трансфицировали в клетки AN3-CA и HL-1 липофектамином 2000 (Thermo Fisher).Через 40 ч при 37 ° C клетки лизировали 25 мМ Tris⋅HCl, pH 7,4, 150 мМ NaCl, 1 мМ этилендиаминтетрауксусной кислоты, 1% (об. / Об.), Nonidet P-40, 5% (об. / Об.). глицерин и 2% (об. / об.) смесь ингибиторов протеазы (Thermo Fisher, 78442). Белки смешивали с образцом буфера с 2% β-меркаптоэтанолом и анализировали электрофорезом в додецилсульфат / полиакриламидном геле и вестерн-блоттингом с использованием антител против FLAG (Sigma, A8592, 1: 5,000) и глицеральдегид-3-фосфатдегидрогеназы (GAPDH) (Multi Наука Биотех, ab011-04, 0.125 мкг / мл).
HESC.
HESC индуцировали к децидуализации в среде DMEM / F12 с 1 мкМ медроксипрогестерона ацетатом (Sigma), 10 нМ эстрадиолом (Cayman Chemical) и 0,5 мМ 8-бром-цАМФ (Sigma). Клетки с обычной средой DMEM / F12 использовали в качестве контроля. В разные моменты времени клетки лизировали, как описано выше. qRT-PCR (описанная ниже) была проведена для анализа уровней мРНК PRL , кодирующего пролактин (маркер децидуализации), и CORIN .Вестерн-блоттинг использовали для анализа эндогенных белков KLF с использованием антител против KLF2 (Abcam, ab203591, 1 мкг / мл) и KLF17 (Abcam, ab84196, 2 мкг / мл). Относительные уровни белка на рентгеновских пленках, полученных с помощью вестерн-блоттинга, определяли количественно с помощью денситометрии. Для иммуногистохимии ГЭСК на покровных стеклах фиксировали 10% параформальдегидом и иммуноокрашивали антителами против Корина (изготовленных в лаборатории) (15), KLF2 (Abcam, ab203591, 5 мкг / мл) и KLF17 (Abcam, ab84196, 5 мкг / мл). мл) или нормальный IgG (Bioworld Technology, BD0051, 2 мкг / мл), а затем вторичное антитело, конъюгированное с пероксидазой хрена (HRP) (поставляется с набором GTVision, Gene Tech, GK5007).После инкубации при 4 ° C в течение ночи с последующей промывкой добавляли раствор субстрата HRP (3,3′-диаминобензидина) и клетки исследовали под микроскопом (Olympus, DP73). Для иммунофлуоресцентного окрашивания использовали вторичные антитела, конъюгированные с Alexa-488 (зеленый) или 594 (красный) (Invitrogen, A21202 и A11012). Ядра клеток окрашивали DAPI (SouthernBiotech) перед конфокальной микроскопией (Olympus, FV1000).
ОТ-ПЦР и qRT-ПЦР.
Суммарные РНК были выделены из культивируемых клеток или тканей мыши с использованием реагента TRIzol (Invitrogen), набора RNeasy (Qiagen) или набора для выделения высокочистой РНК (Omega Bio-tek) для получения комплементарных ДНК первой цепи (кДНК) с кДНК высокой емкости. Набор для обратной транскрипции (Thermo Fisher).ОТ-ПЦР и qRT-ПЦР проводили с олигонуклеотидными праймерами ( SI Приложение , таблица S1) для генов Corin и Klf s мыши или CORIN и PRL человека. Для qRT-PCR реакцию проводили в 20-мкл смеси с 10 мкл LightCycler SYBR Green Master Mix (Roche, 04707516001), матрицами кДНК (17,5 нг) и олигонуклеотидными праймерами (10 пМ каждый) в 40 циклах. Результаты анализировали с помощью программного обеспечения LightCycler 480 (Roche). Относительные уровни мРНК были рассчитаны с использованием значений гена GAPDH или Gapdh в качестве эталона.
ЧИП.
ChIP получали с использованием набора SimpleChIP Plus Enzymatic Chromatin IP (Cell Signaling Technology). Неиндуцированные и индуцированные HESC обрабатывали 1% формальдегидом в течение 10 мин при комнатной температуре и лизировали в буфере, содержащем дитиотреитол и ингибиторы протеаз. Лизаты обрабатывали микрококковой нуклеазой для получения фрагментов хроматина для иммунопреципитации с помощью магнитных шариков и антител против KLF2 (Abcam, ab203591, 1 мкг), KLF9 (Abcam, ab26076, 1 мкг), KLF17 (Abcam, ab84196, 1 мкг) и Гистон 3 (h4) (поставляется с набором ChIP, 10 мкл) (положительный контроль) или нормальный IgG (Bioworld Technology, BD0051, 1 мкг) (отрицательный контроль).Связанные хроматины элюировали с шариков. ДНК разделяли нагреванием, очищали на спин-колонке и использовали для ПЦР и кПЦР с праймерами для сайта KLF в промоторе CORIN (5′-GGA GGA TCT GTC ATT ACG GGT T-3 ‘и 5’-CAA GCT CAA GAG AGA CAA ACT GAA C-3 ‘) или не-KLF сайт (5′-CGG AGG ATC TGT CAT TAC GGG-3′ и 5’-CTG CTC TAC AGA TCC CAC CC-3 ‘).
KLF17 Выбивка в HESC с CRISPR / Cas9.Три пары малых направляющих РНК (sgRNA), нацеленных на ген KLF17 (экзон 1), были разработаны с помощью онлайн-инструментов на веб-сайте CRISPR / Cas9 (https: // wge.stemcell.sanger.ac.uk//find_crisprs). Праймеры — направляющая РНК (пРНК) -K LF17 -1F (5′-CAC CGG CTG AGA TGG AAC AGG AGG CT-3 ‘), gRNA- KLF17 -1R (5′-AAA CAG CCT CCT GTT CCA TCT CAG CC-3 ‘), gRNA- KLF17 -2F (5′-CAC CGG GTC CCC TTT GGT GTC TGT TG -3′), gRNA- KLF17 -2R (5′-AAA CCA ACA GAC ACC AAA GGG GAC CC-3 ‘), gRNA- KLF17 -3F (5′-CAC CGG AGA GAT GCT GGG GTC CCC TT-3′) и gRNA- KLF17 -3R (5′-AAA CAA GGG GAC CCC AGC ATC TCT CC-3 ‘) — отжигали и клонировали в плазмиду PX458 (Addgene), экспрессирующую Cas9-sgRNA, для трансфекции в HESC.Клетки, экспрессирующие зеленый флуоресцентный белок (индикатор), выбранные с помощью проточной цитометрии, выращивали в отдельных колониях после ограничивающего разведения и проверки с помощью ПЦР с последующим секвенированием ДНК и вестерн-блоттингом для нокаута KLF17 .
Исследования на мышах.
Эксперименты с участием мышей были одобрены Комитетом по уходу и использованию животных Университета Сучжоу и проводились в соответствии с руководящими принципами этического обращения и обращения с животными в исследованиях.Мышей содержали в комнатах с регулируемой температурой с 12-часовым / 12-часовым циклом свет-темнота и кормили стандартной диетой для грызунов и неограниченным доступом к воде. Размер выборки был основан на пилотных и опубликованных сопоставимых исследованиях. Исследователи, проводившие эксперименты, не знали генотипов мышей.
Овариэктомия была выполнена анестезированным самкам C57BL / 6 (в возрасте от 8 до 10 недель). Через 9 дней выздоровления мышей случайным образом разделили на три группы для подкожной инъекции кунжутного масла (носитель) (100 мкл), эстрадиола (Cayman Chemical) (50 нг в 100 мкл кунжутного масла) или прогестерона (Sigma) ( 1 мг в 100 мкл кунжутного масла) соответственно.Через 3 дня ежедневной инъекции мышей умерщвляли и выделяли ткани матки для анализа экспрессии генов. Параллельно самок C57BL / 6 (в возрасте от 8 до 10 недель) вязали с самцами C57BL / 6 (в возрасте от 8 до 10 недель) и проверяли на наличие вагинальных пробок. День, когда наблюдалась вагинальная пробка, был определен как G0,5 дня. В разные дни G матки и эмбрионы выделяли для анализа генов, белков и тканей.
Поколение
Klf17 — / — Мышей.Эмбриональные стволовые клетки мыши (идентификатор клона: EPD0099_1_D12) были созданы в Wellcome Trust Sanger Institute, чтобы иметь нарушенный аллель Klf17 ( Klf17 tm1a (KOMP) Wtsi ) с неокассетой и фланкирующими сайтами loxP. экзон 2.Клетки использовали для инъекции бластоцисты для получения химерных и последующих мышей Klf17 + / tm1a на фоне C57BL / 6. Генотипирование проводили с помощью ПЦР с праймерами Klf17 -F (5′-GGT TTC TTT CTG TAA TCC TGG CTA T-3 ‘) и Klf17 -KO-R (5′-CAC AAC GGG TTC TTC TGT TAG TC- 3 ‘) или Klf17 -WT-R (5′-TGC CAT AAC ACT GGT AAA CAA CC-3′). Klf17 + / tm1a мышей были выведены для получения Klf17 + / + , Klf17 + / tm1a и Klf17 tm1a / tm1a дальнейшие анализы. Klf17 + / tm1a и Klf17 tm1a / tm1a мыши обозначаются как Klf17 +/− и Klf17 — / — мыши соответственно , в этом исследовании.
Измерение артериального давления.
Кровяное давление у мышей измеряли с помощью компьютеризированной неинвазивной фотоэлектрической системы «хвост-манжета» (BP-2000, Visitech Systems). Мышей помещали на предварительно нагретую платформу для образцов под крышкой с хвостами в манжете, что позволяло им адаптироваться к устройству и окружающей среде помещения.Каждый тренировочный забег длился не более 30 мин. Через 3 дня акклиматизации, которая проводилась в одно и то же время дня одними и теми же специалистами, было измерено систолическое артериальное давление. Каждое измерение включало 5 циклов предварительной подготовки и 20 регулярных циклов с 5 с между двумя циклами и максимальным давлением в манжете 150 мм рт. Регистрировали средние значения артериального давления.
Мочевые белки.
Образцы мочи были собраны у небеременных и беременных (G12.5 и G18.5) мышей. Уровни белка в моче измеряли с помощью колориметрического анализа Брэдфорда (Nanjing Jiancheng Bioengineering Institute, C035-2) и бычьего сывороточного альбумина в качестве эталонного стандарта.
Гистология тканей, иммуногистохимия и вестерн-блоттинг.
Ткани мыши фиксировали 4% (об. / Об.) Формалином и заливали парафином. Срезы (толщиной 5 мкм) вырезали и помещали на покрытые желатином предметные стекла. Серийные срезы использовали для окрашивания гематоксилином и эозином (H&E), PAS и трихромом Массона или иммуногистохимии на основе опубликованных методов (8, 15). Использовали антитела против цитокератина 18 (Abcam, ab181597, 1,25 мкг / мл) и SMA (Abcam, ab5694, 1 мкг / мл) или нормального IgG (Bioworld Technology, BD0051) (отрицательный контроль).Для обнаружения использовали конъюгированные с HRP вторичные антитела (поставляемые с набором MaxVision, набор-5005, Maxim Biotechnologies). Окрашенные срезы исследовали под микроскопом (Leica DM2000 LED). Для вестерн-блоттинга образцы матки и контрольного сердца и печени гомогенизировали в лизирующем растворе (25). Уровни белка определяли количественно с использованием анализа бицинхониновой кислоты (Thermo Scientific, 23227). Вестерн-блоттинг проводили с использованием первичных антител против Klf17 (Abcam, ab84196, 1: 500) и корина (15) (1: 1000) и вторичного антитела, конъюгированного с HRP (Bioworld Technology, bs13278, 1: 10000).В этих экспериментах рекомбинантный KLF17 и корин, экспрессируемые в клетках HEK293, использовали в качестве положительного контроля. GAPDH использовался в качестве контроля загрузки.
Для иммуногистохимического анализа инвазии трофобластов использовали ткани не менее пяти мышей на группу и не менее двух участков имплантации на мышь. Были подготовлены серийные срезы (> 40 на эмбрион). Положение материнской артерии использовалось в качестве ориентира для ориентации сечения. Стекла, окрашенные цитокератином 18, независимо исследовали два человека.Срезы с наиболее глубокой инвазией трофобластов представлены на рис. 7. Для гистологии клубочков срезы почек окрашивали PAS и трихромом Массона. На окрашенных трихромом срезах Массона эритроциты в клубочках подсчитывали под микроскопом два человека слепым методом. В каждую исследовательскую группу входило не менее трех мышей. Данные были получены по крайней мере из пяти случайно выбранных полей в по крайней мере трех разделах от каждой мыши.
Статистический анализ.
Анализ был выполнен с помощью Prism 7 (Graphpad).Различия анализировали с помощью двустороннего критерия Стьюдента t для сравнения двух групп или ANOVA с последующим апостериорным анализом Тьюки для сравнения трех или более групп. P Значения <0,05 считались статистически значимыми. Данные представлены как среднее значение ± стандартное отклонение или стандартная ошибка среднего, как указано.
Заявление о доступности данных.
Все данные, описанные в статье, представлены в основном тексте SI или приложении .
Благодарности
Мы благодарим Junliang Pan за помощь в анализе промотора гена.Эта работа была частично поддержана грантами Национального фонда естественных наук Китая (81873840, 81671485, 81570457 и 31500636), Национальной программы фундаментальных исследований Китая (2015CB2) и приоритетной академической программы развития институтов высшего образования Цзянсу. Производство мышей поддержано грантом Министерства науки и технологий Китая (2018YFA0801100).
Сноски
Вклад авторов: C.W., Z.W., N.D. и Q.W.спланированное исследование; C.W., Z.W., M.H., T.Z., Y.N., S.S., H.L., C.Z., S.Z. и M.L. проведенное исследование; Y.X. внесены новые реагенты / аналитические инструменты; C.W., Z.W., M.H., T.Z., Y.N., S.S., H.L., M.L., Y.X., N.D. и Q.W. проанализированные данные; и C.W., N.D. и Q.W. написал газету.