Синапс — Википедия
Си́напс (греч. σύναψις, от συνάπτειν — соединение, связь) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём, посредством прохождения ионов из одной клетки в другую.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Однако сам Шеррингтон утверждал, что получил идею этого термина в разговоре от физиолога Майкла Фостера[1].
 Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона
Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексонаПо механизму передачи нервного импульса[править | править код]
- химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
- электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
- смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам[править | править код]
- периферические
- центральные
- аксо-дендритические — с дендритами, в том числе
- аксо-соматические — с телами нейронов;
- аксо-аксональные — между аксонами;
- дендро-дендритические — между дендритами;
 Различные варианты расположения химических синапсов
Различные варианты расположения химических синапсовПо нейромедиатору[править | править код]
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапса[править | править код]
- возбуждающие
- тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса[править | править код]
Типичный синапс — аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидно расширенным окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком плазматической мембраны воспринимающей клетки (в данном случае — участком дендрита).
Между обеими частями имеется синаптическая щель — промежуток шириной 10—50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели — ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый «поцеловал и убежал» (англ. kiss-and-run), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Следствием такой структуры синапса является одностороннее проведение нервного импульса. Существует так называемая синаптическая задержка — время, нужное для передачи нервного импульса. Её длительность составляет около — 0,5 мс.
Так называемый «принцип Дейла» (один нейрон — один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
- В 1897 году Шеррингтон сформулировал представление о синапсах.
- За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль.
- В 1921 австрийский учёный О. Лёви (О. Loewi) установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом (Н. Dale).
- В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче.
- 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
- ↑ French R.D. Some problems and sources in the foundation of modern physiology in Great Britain // Hist. Sci.. — 1971. — № 10. — С. 28-29.
- Савельев А. В. Методология синаптической самоорганизации и проблема дистальных синапсов нейронов // Журнал проблем эволюции открытых систем. — Казахстан, Алматы, 2006. — Т. 8, № 2. — С. 96—104.
- Экклз Д. К. Физиология синапсов. — М.: Мир, 1966. — 397 с.
ru.wikipedia.org
Синаптические связи мозга и верующие нейроны. Тайны мозга. Почему мы во все верим
Синаптические связи мозга и верующие нейроны
Мозг состоит примерно из ста миллиардов нейронов нескольких сотен видов, у каждого из которых есть тело клетки, нисходящий отросток-аксон и многочисленные дендриты и терминали аксона, расходящиеся к другим нейронам и образующие примерно тысячу триллионов синаптических связей между этими ста миллиардами нейронов. Названные цифры ошеломляют. Сто миллиардов нейронов – это 10
Количество синаптических связей мозга равнозначно количеству секунд в 30 миллионах лет.
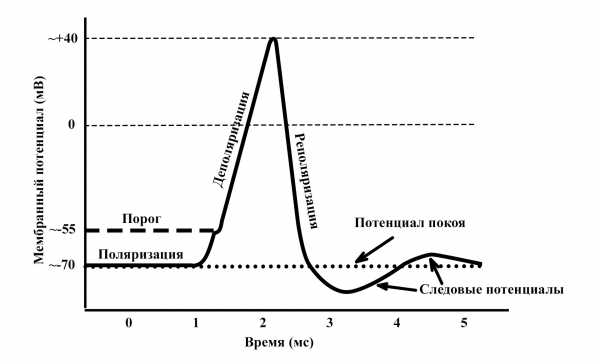
Разумеется, большое количество нейронов обеспечивает значительную вычислительную мощность (как добавление микросхем или карт памяти в компьютер), однако действия производятся в самих отдельных нейронах. Нейронам присуща элегантная простота, вместе с тем это прекрасные в своей сложности машины для обработки электрохимической информации. Внутри нейрона в состоянии покоя больше калия, чем натрия, а преобладание анионов, отрицательно заряженных ионов, создает внутри клетки отрицательный заряд. В зависимости от вида нейрона при введении крошечного электрода в его тело в состоянии покоя мы получим показания –70 мВ (милливольт – одна тысячная вольта). В состоянии покоя клеточная оболочка нейрона непроницаема для натрия, но пропускает калий. При стимуляции нейрона действиями других нейронов (или электрическими манипуляциями любопытных нейробиологов, вооруженных электродами) проницаемость клеточной оболочки меняется, натрий проникает в клетку и таким образом электрический баланс смещается с –70 мВ до нуля. Это явление называется возбудительным постсинаптическим потенциалом, или ВПСП. Синапс – это крохотный зазор между нейронами, следовательно, термин постсинаптический означает, что нейрон на стороне приема сигнала, преодолевающего синаптическую щель, возбуждается, чтобы достичь своего потенциала срабатывания. В отличие от этого, если стимуляция исходит от тормозящего нейрона, напряжение смещается в отрицательную сторону, от –70 мВ до –100 мВ, в итоге срабатывание нейрона становится менее вероятным. Это явление называется тормозящим постсинаптическим потенциалом, или ТПСП. Хотя различных видов нейронов насчитываются сотни, большинство мы можем отнести либо к возбудительным, либо тормозящим по типу действия.
Если при нарастании ВПСП достигает достаточного значения (в результате многочисленных срабатываний одного нейрона за другим или множества связей с другими нейронами), тогда проницаемость клеточной оболочки нейрона достигает критического значения, натрий врывается в него, вызывает мгновенный всплеск напряжения до +50 мВ, оно распространяется по всему телу клетки и поэтапно спускается по аксону в терминали. С той же быстротой напряжение нейрона вновь снижается до –80 мВ, а затем возвращается к –70 мВ в состоянии покоя. Этот процесс приобретения клеточной оболочкой проницаемости для натрия и соответствующего изменения напряжения с отрицательного на положительное, переходящее по аксону к дендритам и синаптическим связям с другими нейронами, называется потенциалом действия. Чаще мы пользуемся выражением «клетка возбудилась». Нарастание ВПСП называется суммацией. Известно два вида: (1) временная суммация, при которой двух ВПСП одного нейрона достаточно для того, чтобы принимающий нейрон достиг критической точки и возбудился; и (2) пространственная суммация, при которой два ВПСП от двух разных нейронов появляются одновременно и их достаточно для того, чтобы принимающий нейрон достиг критической точки и возбудился. Это электрохимическое изменение напряжения происходит стремительно, натриевая проницаемость распространяется последовательно по аксону от тела клетки к терминалям, и это явление, как и следовало ожидать, называется распространением. Скорость распространения зависит от двух условий: (1) диаметра аксона (чем больше, тем быстрее) и (2) миелинизации аксона (чем больше миелиновая оболочка, покрывающая и изолирующая аксон, тем быстрее происходит распространение импульса по нему).[102]
Отметим: если критическая точка возбуждения нейрона не достигнута, он не возбуждается; если критическая точка достигнута, нейрон возбуждается. Эта система работает по принципу «или-или», «все или ничего». Нейроны не возбуждаются «слегка» в ответ на слабые раздражители или «сильно» в ответ на сильные раздражители. Они либо возбуждаются, либо не возбуждаются. Следовательно, нейроны передают информацию одним из трех способов: (1) частотой возбуждения (количеством потенциалов действия в секунду), (2) местом возбуждения (какие именно нейроны возбуждаются) и (3) численностью возбуждения (сколько нейронов возбуждается). Поэтому говорят, что нейроны двоичны по действию, подобны двоичным символам компьютера, 1 и 0, соответствуют сигналу включения или выключения, проходящему или не проходящему по нервному пути. Если рассматривать эти нейронные состояния «включить или выключить» как один из типов ментального состояния, когда один нейрон дает нам два таких состояния (включение или выключение), тогда при обработке информации о мире и управляемом организме у мозга есть 2?1015 возможных вариантов на выбор. Поскольку мы не в состоянии охватить разумом все это число, можно сказать, что мозг во всех отношениях является бесконечно большой машиной для обработки информации.
Каким образом отдельные нейроны и их потенциал действия создают сложные мысли и убеждения? Процесс начинается с так называемого нейронного связывания. «Красный круг» – пример объединения двух входящих сигналов («красный» и «круг») в один воспринимаемый объект, красный круг. Нейронные сигналы от мышц и органов чувств сливаются, двигаясь «вверх по течению», через зоны конвергенции – области мозга, объединяющие информацию, содержащуюся в разных нейронных сигналах (от глаз, ушей, органов осязания и т. д.), чтобы в итоге мы получили представление об объекте в целом, а не о бесчисленных фрагментах изображения. Глядя на перевернутый снимок президента Обамы в главе 4, мы поначалу воспринимаем лицо как одно целое и лишь потом начинаем замечать, что с глазами и ртом что-то не так; как уже объяснялось, причина в том, что две разные нейронные сети действуют с различной скоростью: сначала происходит восприятие лица в целом, затем – деталей этого лица.
Однако связывание – значительно более широкое явление. Объектов, воспринимаемых разными органами чувств, может быть множество, и все они должны связаться воедино в высших областях мозга, чтобы обрести смысл. Крупные отделы мозга, такие, как кора больших полушарий, координируют сигналы от меньших участков мозга, например от височных долей, которые, в свою очередь, объединяют нейронные события от еще меньших компонентов мозга, например от веретенообразной извилины (для распознавания лиц). Это уменьшение происходит на всем пути до уровня единственного нейрона, где нейроны с высокой избирательностью (иногда их называют «бабушкиными») возбуждаются лишь в том случае, когда субъекты видят того, кого знают. Есть нейроны, которые возбуждаются лишь в том случае, когда объект движется слева направо через поле зрения наблюдателя. Есть другие нейроны, которые срабатывают, только когда объект движется справа налево через поле зрения наблюдателя. И есть третьи нейроны, обладающие потенциалом действия только при получении сигналов ВПСП от других нейронов, возбуждающихся в ответ на диагональное движение объектов в поле зрения. Так в нейронных сетях и происходит процесс связывания. Есть даже нейроны, которые возбуждаются, только когда мы видим того, кого узнаем. Нейробиологи из Калтеха Кристоф Кох и Габриэль Крейман совместно с нейрохирургом из Калифорнийского университета в Лос-Анджелесе Ицхаком Фридом обнаружили, например, единственный нейрон, который возбуждается, когда участнику эксперимента показывают снимок Билла Клинтона и более никого. Другой срабатывает, только если участнику показать снимок Дженнифер Энистон, но лишь ее одной, без Брэда Питта.[103]
Разумеется, мы не осознаем работу наших электрохимических систем. Что мы в действительности испытываем, так это субъективные состояния мыслей и чувств, возникающие при объединении нейронных событий и названные философами квалиа. Но даже сами квалиа – один из видов эффекта нейронного связывания, объединения сигналов от бесчисленных нейронных сетей «низшего порядка». Все действительно сводится к электрохимическому процессу нейронного потенциала действия, или к возбуждению нейронов и установлению связи друг с другом с передачей информации. Как им это удается? Опять-таки благодаря химии.
Связь между нейронами возникает в немыслимо крохотной синаптической щели между ними. Когда потенциал действия нейрона устремляется по аксону и достигает его терминалей, он вызывает выброс в синапс мельчайших порций химических трансмиттерных веществ (ХТВ). Полученные соединяющимися нейронами ХТВ действуют как ВПСП, меняя напряжение и проницаемость постсинаптического нейрона, тем самым вызывая его возбуждение и распространение его потенциала действия вниз по аксону до терминалей, где он выбрасывает свои ХТВ в следующий синаптический зазор, и так далее по всей линии нейронной сети. Когда мы ушибаем палец ноги, сигнал боли проходит от болевых рецепторов в тканях нашего пальца ноги весь путь вверх до мозга, который замечает боль и передает сигнал другим участкам мозга, посылающим дополнительные сигналы в сокращающиеся мышцы, чтобы мы отдернули ногу от злополучного препятствия. Все это происходит так быстро, что кажется почти мгновенным.
Существует много видов ХТВ. К самым известным относятся катехоламины, в том числе допамин, норадреналин (норэпинефрин) и адреналин (эпинефрин). ХТВ действуют на постсинаптический нейрон, как ключ на замок. Если ключ подошел и повернулся, нейрон срабатывает; в противном случае дверь остается запертой, а постсинаптический нейрон невозбужденным. После возникновения процесса возбуждения большинство неиспользованных ХТВ возвращается в пресинаптический нейрон, где-либо используется повторно, либо разрушается моноаминоксидазой (МАО) в процессе так называемого первого поглощения. Если в синаптическом зазоре присутствует слишком много ХТВ, тогда остаток всасывается в постсинаптический нейрон в процессе второго поглощения.
Наркотики воздействуют на синапсы, выброс ХТВ и последующие процессы поглощения. Например, амфетамины ускоряют выброс ХТВ в синапсы, тем самым ускоряя процесс нейронной коммуникации, потому и называются speed («скорость»). Резерпин, который некогда был обычным назначением при психозах, разрушает пузырьки с ХТВ в пресинаптическом нейроне, поэтому МАО уничтожают их еще до использования, в итоге замедляют работу нейронных сетей, контролируют маниакальные состояния, гипертензию и другие симптомы гиперактивности нервной системы. Кокаин блокирует первое поглощение, поэтому ХТВ просто задерживаются в синапсе и способствуют ускоренному возбуждению нейронов, доводят нейронные сети до состояния взвинченности – вспомните Робина Уильямса с микрофоном перед аудиторией; в сущности, сам Уильямс в значительной мере приписывает успех своих комедий в 1980-х годах собственной кокаиновой зависимости. Как один из самых распространенных ХТВ, допамин играет решающую роль в беспрепятственной коммуникации между нейронами и мышцами, а когда его недостаточно, у пациентов наблюдаются потеря регуляции моторики и неудержимая дрожь. Эти проявления называются болезнью Паркинсона, один из методов лечения которой – L-dopa, агонист допамина, стимулирующий его выработку.
Как нам построить всю систему снизу доверху, начиная с химических трансмиттерных веществ, таких как допамин, и связывая сигналы в единую систему убеждений? Посредством поведения. Напомню, что первичная функция мозга – управлять телом и помогать ему выжить. Один из способов сделать это – посредством ассоциативного обучения, или паттерничности. Это и есть связующее звено между нейронным потенциалом действия и человеческими поступками.
Поделитесь на страничкеСледующая глава >
psy.wikireading.ru
как наш мозг создает нейронные связи и формирует привычки и интеллект — T&P
Гормоны влияют на механизмы образования эмоций и действие различных нейрохимических веществ, и, как следствие, участвуют в формировании устойчивых привычек. Автор книги «Гормоны счастья» заслуженный профессор Калифорнийского университета Лоретта Грациано Бройнинг предлагает пересмотреть шаблоны нашего поведения и научиться запускать действие серотонина, дофамина, эндорфина и окситоцина. T&P публикуют главу из книги о том, как самонастраивается наш мозг, реагируя на опыт и формируя соответствующие нейронные связи.
Лоретта Грациано Бройнинг
основатель Inner Mammal Institute, заслуженный профессор Калифорнийского университета, автор нескольких книг, ведет блог «Your Neurochemical Self» на сайте PsychologyToday.com
Перекладывая нейронные пути
Каждый человек рождается с множеством нейронов, но очень небольшим количеством связей между ними. Эти связи строятся по мере взаимодействия с окружающим нас миром и в конечном счете и создают нас такими, какие мы есть. Но иногда у вас возникает желание несколько модифицировать эти сформировавшиеся связи. Казалось бы, это должно быть легко, потому что они сложились у нас без особых усилий с нашей стороны еще в молодости. Однако формирование новых нейронных путей во взрослом возрасте оказывается неожиданно сложным делом. Старые связи настолько эффективны, что отказ от них создает у вас ощущение, что возникает угроза выживанию. Любые новые нервные цепочки являются весьма хрупкими по сравнению со старыми. Когда вы сможете понять, как трудно создаются в мозгу человека новые нейронные пути, вы будете радоваться своей настойчивости в этом направлении больше, чем ругать себя за медленный прогресс в их формировании.
Пять способов, с помощью которых самонастраивается ваш мозг
Мы, млекопитающие, способны в течение жизни создавать нейронные связи, в отличие от видов с устойчивыми связями. Эти связи создаются по мере того, как окружающий нас мир воздействует на наши органы чувств, которые посылают соответствующие электрические импульсы в мозг. Эти импульсы прокладывают нейронные пути, по которым в будущем быстрее и легче побегут другие импульсы. Мозг каждого отдельного человека настроен на индивидуальный опыт. Ниже приведены пять способов, с помощью которых опыт физически меняет ваш мозг.
Жизненный опыт изолирует молодые нейроны
Постоянно работающий нейрон с течением времени покрывается оболочкой из особого вещества, которое называется миелин. Это вещество значительно повышает эффективность нейрона как проводника электрических импульсов. Это можно сравнить с тем, что изолированные провода могут выдерживать значительно большую нагрузку, чем оголенные. Покрытые миелиновой оболочкой нейроны работают без затраты излишних усилий, что свойственно медленным, «открытым» нейронам. Нейроны с миелиновой оболочкой выглядят скорее белыми, чем серыми, поэтому мы разделяем наше мозговое вещество на «белое» и «серое».
В основном покрытие нейронов миелином завершается у ребенка к возрасту двух лет, по мере того как его тело научается двигаться, видеть и слышать. Когда рождается млекопитающее, в его мозгу должна сформироваться ментальная модель окружающего его мира, что предоставит ему возможности для выживания. Поэтому выработка миелина у ребенка максимальна при рождении, а к семи годам она несколько снижается. К этому времени вам уже не надо учить заново истины, что огонь обжигает, а земное тяготение может заставить вас упасть.
Если вы думаете, что миелин «зря расходуется» на усиление нейронных связей именно у молодых, то следует понимать, что природа устроила именно так по обоснованным эволюционным причинам. На протяжении большей части истории человечества люди заводили детей сразу по достижении половой зрелости. Нашим предкам нужно было успеть решить первоочередные насущные задачи, которые обеспечивали выживание их потомства. Во взрослом состоянии они больше использовали новые нейронные связи, чем перенастраивали старые.С достижением периода полового созревания человека формирование миелина в его организме вновь активизируется. Это происходит из-за того, что млекопитающему предстоит осуществить новую настройку своего мозга на поиск наилучшего брачного партнера. Часто в период спаривания животные мигрируют в новые группы. Поэтому им приходится привыкать к новым местам в поисках пищи, а также к новым соплеменникам. В поисках брачной пары люди также нередко вынуждены перемещаться в новые племена или кланы и постигать новые обычаи и культуру. Рост выработки миелина в период полового созревания как раз всему этому и способствует. Естественный отбор устроил мозг таким, что именно в этот период он меняет ментальную модель окружающего мира.
Все, что вы целенаправленно и постоянно делаете в годы своего «миелинового расцвета», создает мощные и разветвленные нейронные пути в вашем мозгу. Именно поэтому так часто гениальность человека проявляется именно в детстве. Именно поэтому маленькие горнолыжники так лихо пролетают мимо вас на горных спусках, которые вы не можете освоить, сколько ни стараетесь. Именно поэтому таким трудным становится изучение иностранных языков с окончанием юношеского возраста. Будучи уже взрослыми, вы можете запоминать иностранные слова, но чаще всего вы не можете быстро подбирать их для выражения своих мыслей. Это происходит потому, что вербальная память концентрируется у вас в тонких, не покрытых миелином нейронах. Мощные миелинизированные нейронные связи заняты у вас высокой мыслительной деятельностью, поэтому новые электрические импульсы с трудом находят свободные нейроны. […]
Колебания активности организма в миелинизации нейронов могут помочь вам понять, почему у людей возникают те или иные проблемы в разные периоды жизни. […] Помните, что человеческий мозг не достигает своей зрелости автоматически. Поэтому часто говорят, что мозг у подростков еще не вполне сформировавшийся. Мозг «миелинирует» весь наш жизненный опыт. Так что если в жизни подростка будут иметь место эпизоды, когда он получает незаслуженное вознаграждение, то он накрепко запоминает, что награду можно получить и без усилий. Некоторые родители прощают подросткам плохое поведение, говоря, что «их мозг еще не полностью оформился». Именно поэтому очень важно целенаправленно контролировать тот жизненный опыт, который они впитывают. Если позволить подростку избегать ответственности за свои действия, то можно сформировать у него разум, который будет ожидать возможности уклонения от такой ответственности и в дальнейшем. […]
Жизненный опыт повышает эффективность работы синапса
Синапс — это место контакта (небольшой промежуток) между двумя нейронами. Электрический импульс в нашем мозгу может передвигаться только при том условии, что он достигает конца нейрона с достаточной силой, чтобы «перепрыгнуть» через этот промежуток к следующему нейрону. Эти барьеры помогают нам фильтровать на самом деле важную входящую информацию от не имеющего значения так называемого «шума». Прохождение электрического импульса через синаптические промежутки — это очень сложный природный механизм. Его можно представить себе так, что на кончике одного нейрона скапливается целая флотилия лодок, которая транспортирует нейронную «искру» в специальные приемные доки, имеющиеся у рядом расположенного нейрона. С каждым разом лодки лучше справляются с транспортировкой. Вот почему получаемый нами опыт увеличивает шансы передачи электрических сигналов между нейронами. В мозге человека имеется более 100 триллионов синаптических связей. И наш жизненный опыт играет важную роль, чтобы проводить по ним нервные импульсы так, чтобы это соответствовало интересам выживания.
На сознательном уровне вы не можете решать, какие именно синаптические связи вам следует развивать. Они формируются двумя основными способами:
1) Постепенно, путем многократного повторения.
2) Одномоментно, под воздействием сильных эмоций.
[…] Синаптические связи строятся на основе повторения или эмоций, пережитых вами в прошлом. Ваш разум существует за счет того, что ваши нейроны образовали связи, которые отражают удачный и неудачный опыт. Некоторые эпизоды из этого опыта были «закачаны» в ваш мозг благодаря «молекулам радости» или «молекулам стресса», другие были закреплены в нем благодаря постоянным повторениям. Когда модель окружающего мира соответствует той информации, которая содержится в ваших синаптических связях, электрические импульсы пробегают по ним легко, и вам кажется, что вы вполне в курсе происходящих вокруг вас событий.
Нейронные цепочки формируются только за счет активных нейронов
Те нейроны, которые активно не используются мозгом, начинают постепенно ослабевать уже у двухлетнего ребенка. Как ни странно, это способствует развитию его интеллекта. Сокращение числа активных нейронов позволяет малышу не скользить рассеянным взглядом по всему вокруг, что свойственно новорожденному, а опираться на нейронные пути, которые у него уже сформировались. Двухлетний малыш способен уже самостоятельно концентрироваться на том, что доставляло ему в прошлом приятные ощущения типа знакомого лица или бутылочки с его любимой едой. Он может остерегаться того, что в прошлом вызвало у него отрицательные эмоции, например драчливый товарищ по играм или закрытая дверь. Юный мозг полагается уже на свой небольшой жизненный опыт в том, что касается удовлетворения нужд и избегания потенциальных угроз.
Как бы ни строились нейронные связи в мозге, вы ощущаете их как «истину»
В возрасте от двух до семи лет процесс оптимизации мозга у ребенка продолжается. Это заставляет его соотносить новый опыт со старым, вместо того чтобы накапливать новые переживания каким-то отдельным блоком. Тесно переплетенные нейронные связи и нервные пути составляют основу нашего интеллекта. Мы создаем их, разветвляя старые нейронные «стволы», вместо того чтобы создавать новые. Таким образом, к семи годам мы обычно четко видим то, что уже однажды видели, и слышим уже однажды услышанное.
Вы можете подумать, что это плохо. Однако подумайте над ценностью всего этого. Представьте себе, что вы солгали шестилетнему ребенку. Он верит вам, потому что его мозг жадно впитывает все, что ему предлагается. Теперь предположите, что вы обманули ребенка восьми лет. Он уже подвергает ваши слова сомнению, потому что сравнивает поступающую информацию с уже имеющейся у него, а не просто «проглатывает» новые сведения. В возрасте восьми лет ребенку уже труднее формировать новые нейронные связи, что толкает его на использование уже имеющихся. Опора на старые нейронные цепочки позволяет ему распознать ложь. Это имело огромное значение с точки зрения выживания для того времени, когда родители умирали молодыми и детям с малых лет приходилось привыкать заботиться о себе. В юные годы мы формируем определенные нейронные связи, позволяя другим постепенно угасать. Некоторые из них исчезают, как ветер уносит осенние листья. Это помогает сделать мыслительный процесс человека более эффективным и целенаправленным. Конечно, с возрастом вы получаете все новые знания. Однако эта новая информация концентрируется в тех областях мозга, в которых уже существуют активные электрические пути. Например, если наши предки рождались в охотничьих племенах, то быстро набирали опыт охотника, а если в племенах землепашцев — сельскохозяйственный опыт. Таким образом мозг настраивался на выживание в том мире, в котором они реально существовали. […]
Между активно используемыми вами нейронами образуются новые синаптические связи
Каждый нейрон может иметь много синапсисов, потому что у него бывает много отростков или дендритов. Новые отростки у нейронов образуются при его активной стимуляции электроимпульсами. По мере того как дендриты растут в направлении точек электрической активности, они могут приблизиться настолько, что электрический импульс от других нейронов может преодолеть расстояние между ними. Таким образом рождаются новые синаптические связи. Когда подобное происходит, на уровне сознания вы получаете связь между двумя идеями, например.
Свои синаптические связи вы ощущать не можете, но легко можете увидеть это в других. Человек, любящий собак, смотрит на весь окружающий мир через призму этой привязанности. Человек, увлеченный современными технологиями, все на свете связывает с ними. Любитель политики оценивает окружающую реальность политически, а религиозно убежденный человек — с позиций религии. Один человек видит мир позитивно, другой — негативно. Как бы ни строились нейронные связи в мозге, вы не ощущаете их как многочисленные отростки, похожие на щупальца осьминога. Вы ощущаете эти связи как «истину».
Рецепторы эмоций развиваются или атрофируются
Для того чтобы электрический импульс мог пересечь синаптическую щель, дендрит с одной стороны должен выбросить химические молекулы, которые улавливаются специальными рецепторами другого нейрона. Каждое из нейрохимических веществ, вырабатываемых нашим мозгом, имеет сложную структуру, которая воспринимается только одним специфическим рецептором. Она подходит к рецептору, как ключ к замку. Когда вас захлестывают эмоции, то вырабатывается больше нейрохимических веществ, чем может уловить и обработать рецептор. Вы чувствуете себя ошеломленным и дезориентированным до тех пор, пока ваш мозг не создаст больше рецепторов. Так вы адаптируетесь к тому, что «вокруг вас что-то происходит».
Когда рецептор нейрона продолжительное время неактивен, он исчезает, оставляя место для появления других рецепторов, которые могут вам понадобиться. Гибкость в природе означает, что рецепторы у нейронов должны либо использоваться, либо они могут потеряться. «Гормоны радости» постоянно присутствуют в мозге, осуществляя поиск «своих» рецепторов. Именно так вы и «узнаете» причину своих позитивных ощущений. Нейрон «срабатывает», потому что подходящие молекулы гормонов открывают замок его рецептора. А затем на основе этого нейрона создается целая нейронная цепь, которая подсказывает вам, откуда ожидать радости в будущем.
Изображения: © iStock.
theoryandpractice.ru
Нейроны и синапсы или как работает мозг | krok8.com
Содержание:Основным компонентом нервной системы в целом и мозга в частности является нейрон или нервная клетка, «клетки мозга». Нейрон – это электрически возбудимая клетка, которая обрабатывает и передает информацию посредством электрохимической сигнализации.
В отличие от других клеток, нейроны никогда не делятся и не отмирают, чтобы их заменили новые. По той же причине они обычно не могут быть восстановлены после потери, хотя есть несколько исключений.
В отличие от других клеток организма, большинство нейронов в человеческом мозге способны делиться только для того, чтобы создавать новые клетки (процесс, называемый нейрогенезом) во время развития плода и в течение нескольких месяцев после рождения.
Эти клетки мозга могут увеличиваться в размерах до возраста около восемнадцати лет, но они, по существу, рассчитаны на всю жизнь.
Интересно, что единственной областью мозга, где нейрогенез, как было показано, продолжается на протяжении всей жизни, является гиппокамп, область, необходимая для кодирования и хранения памяти.
Объем памяти человека
Средний человеческий мозг имеет около 100 миллиардов нейронов (или нервных клеток) и нейроглии (или глиальные клетки), которые служат для поддержки и защиты нейронов.
Каждый нейрон может быть связан с 10 000 других нейронов, передавая сигналы друг другу через 1000 триллионов синаптических соединений, что, по некоторым оценкам, эквивалентно компьютеру с процессором со скоростью 1 триллион бит в секунду.
Оценки объема памяти человеческого мозга сильно варьируются от 1 до 1000 терабайт (для сравнения, 19 миллионов томов в Библиотеке Конгресса США представляют около 10 терабайт данных).
Обзор механизмов и принципов передачи информации в мозге, работа памяти человека.
Передача информации в мозге, например, во время процессов кодирования и извлечения памяти, достигается с помощью комбинации химических веществ и электричества. Это очень сложный процесс, включающий множество взаимосвязанных этапов, но краткий обзор можно произвести.
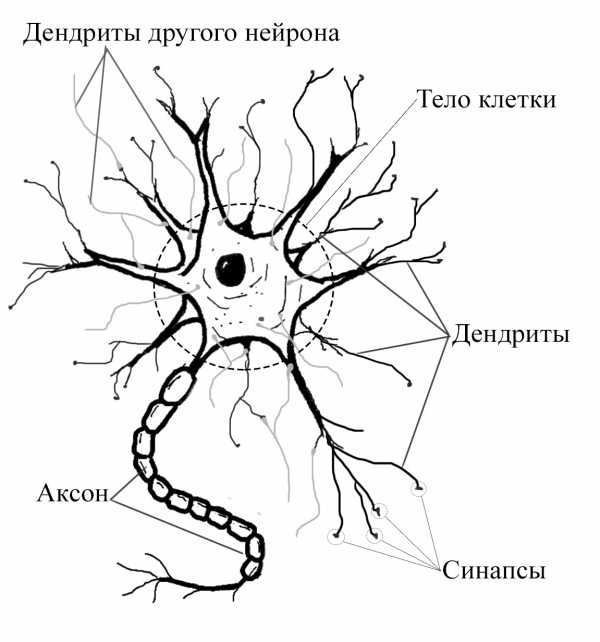
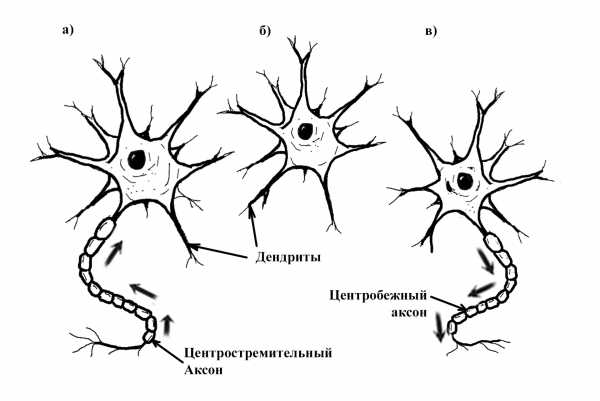 Схема нейрона. Изображение из Википедии
Схема нейрона. Изображение из ВикипедииТипичный нейрон обладает сомой (клеточным телом содержащим клеточное ядро), дендритами (Дендрит — ветвящийся отросток нервной клетки (нейрона), воспринимающий сигналы от других нейронов, рецепторных клеток или непосредственно от внешних раздражителей.) (разветвлёнными отростками, прикрепленными к клеточному телу в сложном ветвящемся «дендритном дереве») и одним аксоном (Аксон — это нейрит (длинный цилиндрический отросток нервной клетки), по которому нервные импульсы идут от тела клетки (сомы) к иннервируемым органам и другим нервным клеткам.) (длинным цилиндрическим отростком, который может быть в тысячи раз длиннее сомы).
Каждый нейрон поддерживает градиент напряжения на своей мембране из-за метаболически обусловленных различий в ионах натрия, калия, хлорида и кальция внутри клетки, каждый из которых имеет различный заряд.
Если напряжение существенно изменяется, генерируется электрохимический импульс, называемый потенциалом действия (или нервным импульсом). Эта электрическая активность может быть измерена и отображена в виде волновой формы, называемой мозговой волной или ритмом мозга.
Этот импульс быстро распространяется по аксону клетки и передается через специализированное соединение, известное как синапс (Синапс — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой.) , к соседнему нейрону, который получает его через свои дендриты.
Синапс представляет собой сложное мембранное соединение или разрыв (фактический разрыв, также известный как синаптическая щель, составляет порядка 20 нанометров, или 20 миллионных миллиметра), используется для передачи сигналов между клетками, и поэтому известен как синаптическая связь.
Хотя синаптические связи аксон-дендрит являются нормой, возможны и другие варианты (например, дендрит-дендрит, аксон-аксон, дендрит-аксон). Типичный нейрон срабатывает 5 – 50 раз каждую секунду.
Таким образом, каждый отдельный нейрон может образовывать тысячи связей с другими нейронами, давая мозгу более 100 триллионов синапсов (до 1000 триллионов, по некоторым оценкам).
Функционально связанные нейроны соединяются друг с другом, образуя нейронные сети. Однако связи между нейронами не статичны, они меняются со временем.
Чем больше сигналов посылается между двумя нейронами, тем сильнее растет связь, и поэтому с каждым новым опытом и каждым запоминающимся событием или фактом мозг слегка перестраивает свою физическую структуру.
В детстве, и особенно в подростковом возрасте, происходит процесс, известный как «синаптическая обрезка».
Несмотря на то, что мозг продолжает расти и развиваться, общее количество нейронов и синапсов сокращается до 50%, удаляя ненужные нейронные структуры и позволяя им заменяться более сложными и эффективными структурами, более подходящими к требований взрослой жизни.
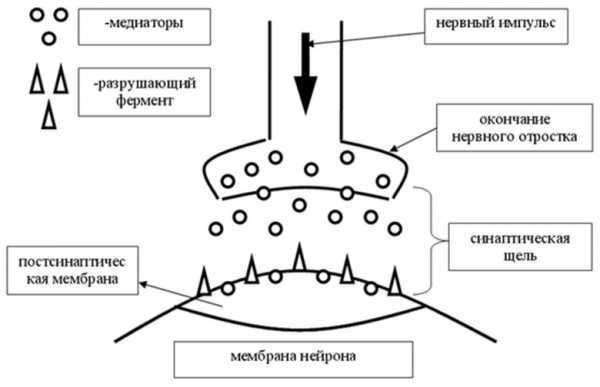 Синаптическая передача. Изображение из Википедии
Синаптическая передача. Изображение из ВикипедииВзаимодействие нейронов не только электрическое, но и электрохимическое. Каждый аксонный терминал содержит тысячи связанных мембраной мешочков, называемых везикулами (Синаптические везикулы (или синаптические пузырьки) находятся в пресинаптических границах в нейронах и складируют нейромедиаторы.) , которые, в свою очередь, содержат тысячи молекул нейротрансмиттеров (Нейромедиаторы (нейротрансмиттеры, посредники, «медиаторы») — биологически активные химические вещества, посредством которых осуществляется передача электрохимического импульса от нервной клетки через синаптическое пространство между нейронами, а также, например, от нейронов к мышечной ткани или железистым клеткам.) .
Нейротрансмиттеры – это химические посыльные которые передают, усиливают и модулируют сигналы между нейронами и другими клетками.
- Двумя наиболее распространенными нейротрансмиттерами в мозге являются аминокислоты глутамат и ГАМК;
- другими важными нейротрансмиттерами являются ацетилхолин, допамин, адреналин, гистамин, серотонин и мелатонин.
При стимуляции электрическим импульсом высвобождаются нейромедиаторы различных типов и пересекают клеточную мембрану в синаптическую щель между нейронами.
Эти химические вещества затем связываются с химическими рецепторами в дендритах принимающего (постсинаптического) нейрона.
В процессе они вызывают изменения проницаемости клеточной мембраны для конкретных ионов, открывая специальные ворота или каналы, которые впускают поток заряженных частиц (ионы кальция, натрия, калия и хлорида).
Это влияет на потенциальный заряд принимающего нейрона, который затем запускает новый электрический сигнал в принимающем нейроне. Весь процесс занимает менее одной пятисотой секунды.
Таким образом, сообщение в мозгу преобразуется, когда оно перемещается от одного нейрона к другому, от электрического сигнала к химическому сигналу и обратно, в непрерывную цепь событий, которая является основой всей деятельности мозга.
Электрохимический сигнал выпущенный определенным нейротрансмиттером может быть как стимулирующим (например, ацетилхолин, глутамат, аспартат, норадреналин, гистамин), так и ингибирующим (например, ГАМК, глицин, сератонин), а некоторые (например, дофамин) могут оказывать и то и другое действие.
Тонкие вариации в механизмах нейромедиации позволяют мозгу реагировать на различные требования, предъявляемые к нему, включая кодирование, консолидацию, хранение и извлечение воспоминаний.
Общие сведения о глиальных клетках (нейроглия, глия)
Как уже упоминалось, помимо нейронов, мозг содержит примерно равную массу глиальных клеток (нейроглия или просто глия), наиболее распространенными типами которых являются олигодендроциты, астроциты (Астроцит — тип нейроглиальной клетки звездчатой формы с многочисленными отростками.) и микроглии.
Поскольку они намного меньше, чем нейроны, их в 10 раз больше, а различные области мозга имеют более высокую или более низкую концентрацию глий.
Раньше считалось, что роль глиальных (Нейроглия (глия) — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов.) клеток ограничивается физической поддержкой, питанием и восстановлением нейронов центральной нервной системы.
Тем не менее, более недавние исследования показывают, что глия, особенно астроциты, на самом деле выполняют гораздо более активную роль в коммуникации мозга и нейропластичности, хотя степень и механизм этой роли все еще неопределенны, и значительный объем современных исследований мозга в настоящее время сосредоточен на глиальных клетках.
Источники:- NEURONS & SYNAPSES
Все материалы носят ознакомительный характер. [Отказ от ответственности krok8.com]
krok8.com
В человеческом мозге столько же «транзисторов», сколько в мировой ИТ-инфраструктуре
Стэнфордские нейробиологи потратили несколько лет, разрабатывая новый способ 3D-сканирования мозга. Они совместили объёмную компьютерную томографию (array tomography — техника «антенных решёток» из радиоастрономии) и специально разработанный софт, чтобы получить объёмную и реалистичную 3D-модель. Такую, по которой можно перемещаться, масштабировать и вращать её в разных измерениях.Сканирование от мягкой оболочки коры мозга мыши через шесть слоёв и подкорковое белое вещество к прилегающему полосатому телу.
Изучив полученную картину, учёные пришли к выводу, что синапсы устроены гораздо сложнее, чем предполагалось раньше.
Здоровый человеческий мозг содержит около 200 млрд нервных клеток, которые соединяются друг с другом сотнями триллионов синапсов. От каждой нервной клетки могут отходить десятки тысяч синапсов. В одной только коре больших полушарий человека находится около 125 трлн синапсов — в 1500 раз больше, чем звёзд в нашей галактике.
По результатам визуальной реконструкции данных учёные обнаружили, что каждый синапс содержит около 1000 молекулярных «переключателей», эдаких аналоговых транзисторов. То есть отдельный синапс можно сравнить с микропроцессором. Получается, что количество «транзисторов» в человеческом мозге теперь нужно увеличить на три порядка. Их больше, чем транзисторов во всех компьютерах на планете и маршрутизаторах вместе взятых.
Расчёт такой:
- 1,25E14 синапсов в мозге
- в каждом синапсе 1000 молекулярных переключателей
- получается 1,25E17 молекулярных переключателей в мозге
- с 2005 года в микропроцессорах примерно 1E8 транзисторов (с 2010 года начали делать 1E9)
- предположим, что в мире 1E9 компьютеров
- получается, что во всех компьютерах на Земле 1E17 транзисторов
- 1,25E17 > 1E17, ч.т.д.
С одной стороны, кто-то может сделать вывод, что мы ещё очень далеки от создания искусственного интеллекта. С другой стороны, уже на базе текущей инфраструктуры можно запустить реальную модель человеческого мозга 1:1. Нужно только понять, по какой программе работают синапсы, то есть расшифровать всю биохимию.
Визуальная реконструкция данных объёмной компьютерной томографии синапсов в постцентральной извилине мыши, которая реагирует на стимуляцию волоском. Нейроны подсвечиваются введённым в мозг флуоресцентным красителем зелёного цвета. За счёт этого в массе нервных клеток можно различить более мелкие синапсы.
Результаты исследования по визуальной реконструкции данных с объёмной компьютерной томографии опубликованы в журнале Neuron. Учёные получили патент и подали заявку на второй, а также основали компанию, чтобы коммерциализировать новый метод томографии.
via Health Tech
habr.com
Синаптические связи мозга и верующие нейроны
Мозг состоит примерно из ста миллиардов нейронов нескольких сотен видов, у каждого из которых есть тело клетки, нисходящий отросток-аксон и многочисленные дендриты и терминали аксона, расходящиеся к другим нейронам и образующие примерно тысячу триллионов синаптических связей между этими ста миллиардами нейронов. Названные цифры ошеломляют. Сто миллиардов нейронов – это 1011, или единица, а за ней 11 нулей: 100000000000. Связи тысячи триллионов – квадрильон, или 1015, или единица, за которой следуют 15 нулей: 1000000000000000. Нейронов в человеческом мозге примерно столько же, сколько звезд в галактике Млечный Путь – в буквальном смысле слова астрономическое число! Количество синаптических связей мозга равнозначно количеству секунд в 30 миллионах лет. Задумайтесь об этом на минуту. Начните отсчитывать секунды способом «одна одна тысяча, две одна тысяча, три одна тысяча…» Когда доберетесь до 86400, получится количество секунд в сутках, когда достигнете 31536000, – количество секунд в году, когда наконец дойдете до одного триллиона секунд, значит, вы считаете уже примерно 30 тысяч лет. А теперь повторите этот счет продолжительностью в 30 тысяч лет еще одну тысячу раз, и вы отсчитаете количество синаптических связей в своем мозге.
Количество синаптических связей мозга равнозначно количеству секунд в 30 миллионах лет.
Разумеется, большое количество нейронов обеспечивает значительную вычислительную мощность (как добавление микросхем или карт памяти в компьютер), однако действия производятся в самих отдельных нейронах. Нейронам присуща элегантная простота, вместе с тем это прекрасные в своей сложности машины для обработки электрохимической информации. Внутри нейрона в состоянии покоя больше калия, чем натрия, а преобладание анионов, отрицательно заряженных ионов, создает внутри клетки отрицательный заряд. В зависимости от вида нейрона при введении крошечного электрода в его тело в состоянии покоя мы получим показания –70 мВ (милливольт – одна тысячная вольта). В состоянии покоя клеточная оболочка нейрона непроницаема для натрия, но пропускает калий. При стимуляции нейрона действиями других нейронов (или электрическими манипуляциями любопытных нейробиологов, вооруженных электродами) проницаемость клеточной оболочки меняется, натрий проникает в клетку и таким образом электрический баланс смещается с –70 мВ до нуля. Это явление называется возбудительным постсинаптическим потенциалом , или ВПСП. Синапс – это крохотный зазор между нейронами, следовательно, термин постсинаптический означает, что нейрон на стороне приема сигнала, преодолевающего синаптическую щель, возбуждается, чтобы достичь своего потенциала срабатывания. В отличие от этого, если стимуляция исходит от тормозящего нейрона, напряжение смещается в отрицательную сторону, от –70 мВ до –100 мВ, в итоге срабатывание нейрона становится менее вероятным. Это явление называется тормозящим постсинаптическим потенциалом , или ТПСП. Хотя различных видов нейронов насчитываются сотни, большинство мы можем отнести либо к возбудительным, либо тормозящим по типу действия. Если при нарастании ВПСП достигает достаточного значения (в результате многочисленных срабатываний одного нейрона за другим или множества связей с другими нейронами), тогда проницаемость клеточной оболочки нейрона достигает критического значения , натрий врывается в него, вызывает мгновенный всплеск напряжения до +50 мВ, оно распространяется по всему телу клетки и поэтапно спускается по аксону в терминали. С той же быстротой напряжение нейрона вновь снижается до –80 мВ, а затем возвращается к –70 мВ в состоянии покоя. Этот процесс приобретения клеточной оболочкой проницаемости для натрия и соответствующего изменения напряжения с отрицательного на положительное, переходящее по аксону к дендритам и синаптическим связям с другими нейронами, называется потенциалом действия . Чаще мы пользуемся выражением «клетка возбудилась». Нарастание ВПСП называется суммацией . Известно два вида: (1) временная суммация , при которой двух ВПСП одного нейрона достаточно для того, чтобы принимающий нейрон достиг критической точки и возбудился; и (2) пространственная суммация , при которой два ВПСП от двух разных нейронов появляются одновременно и их достаточно для того, чтобы принимающий нейрон достиг критической точки и возбудился. Это электрохимическое изменение напряжения происходит стремительно, натриевая проницаемость распространяется последовательно по аксону от тела клетки к терминалям, и это явление, как и следовало ожидать, называется распространением . Скорость распространения зависит от двух условий: (1) диаметра аксона (чем больше, тем быстрее) и (2) миелинизации аксона (чем больше миелиновая оболочка, покрывающая и изолирующая аксон, тем быстрее происходит распространение импульса по нему).[102] Отметим: если критическая точка возбуждения нейрона не достигнута, он не возбуждается; если критическая точка достигнута, нейрон возбуждается. Эта система работает по принципу «или-или», «все или ничего». Нейроны не возбуждаются «слегка» в ответ на слабые раздражители или «сильно» в ответ на сильные раздражители. Они либо возбуждаются, либо не возбуждаются. Следовательно, нейроны передают информацию одним из трех способов: (1) частотой возбуждения (количеством потенциалов действия в секунду), (2) местом возбуждения (какие именно нейроны возбуждаются) и (3) численностью возбуждения (сколько нейронов возбуждается). Поэтому говорят, что нейроны двоичны по действию, подобны двоичным символам компьютера, 1 и 0, соответствуют сигналу включения или выключения, проходящему или не проходящему по нервному пути. Если рассматривать эти нейронные состояния «включить или выключить» как один из типов ментального состояния, когда один нейрон дает нам два таких состояния (включение или выключение), тогда при обработке информации о мире и управляемом организме у мозга есть 2×1015 возможных вариантов на выбор. Поскольку мы не в состоянии охватить разумом все это число, можно сказать, что мозг во всех отношениях является бесконечно большой машиной для обработки информации. Каким образом отдельные нейроны и их потенциал действия создают сложные мысли и убеждения? Процесс начинается с так называемого нейронного связывания . «Красный круг» – пример объединения двух входящих сигналов («красный» и «круг») в один воспринимаемый объект, красный круг. Нейронные сигналы от мышц и органов чувств сливаются, двигаясь «вверх по течению», через зоны конвергенции – области мозга, объединяющие информацию, содержащуюся в разных нейронных сигналах (от глаз, ушей, органов осязания и т. д.), чтобы в итоге мы получили представление об объекте в целом, а не о бесчисленных фрагментах изображения. Глядя на перевернутый снимок президента Обамы в главе 4, мы поначалу воспринимаем лицо как одно целое и лишь потом начинаем замечать, что с глазами и ртом что-то не так; как уже объяснялось, причина в том, что две разные нейронные сети действуют с различной скоростью: сначала происходит восприятие лица в целом, затем – деталей этого лица. Однако связывание – значительно более широкое явление. Объектов, воспринимаемых разными органами чувств, может быть множество, и все они должны связаться воедино в высших областях мозга, чтобы обрести смысл. Крупные отделы мозга, такие, как кора больших полушарий, координируют сигналы от меньших участков мозга, например от височных долей, которые, в свою очередь, объединяют нейронные события от еще меньших компонентов мозга, например от веретенообразной извилины (для распознавания лиц). Это уменьшение происходит на всем пути до уровня единственного нейрона, где нейроны с высокой избирательностью (иногда их называют «бабушкиными») возбуждаются лишь в том случае, когда субъекты видят того, кого знают. Есть нейроны, которые возбуждаются лишь в том случае, когда объект движется слева направо через поле зрения наблюдателя. Есть другие нейроны, которые срабатывают, только когда объект движется справа налево через поле зрения наблюдателя. И есть третьи нейроны, обладающие потенциалом действия только при получении сигналов ВПСП от других нейронов, возбуждающихся в ответ на диагональное движение объектов в поле зрения. Так в нейронных сетях и происходит процесс связывания. Есть даже нейроны, которые возбуждаются, только когда мы видим того, кого узнаем. Нейробиологи из Калтеха Кристоф Кох и Габриэль Крейман совместно с нейрохирургом из Калифорнийского университета в Лос-Анджелесе Ицхаком Фридом обнаружили, например, единственный нейрон, который возбуждается, когда участнику эксперимента показывают снимок Билла Клинтона и более никого. Другой срабатывает, только если участнику показать снимок Дженнифер Энистон, но лишь ее одной, без Брэда Питта.[103] Разумеется, мы не осознаем работу наших электрохимических систем. Что мы в действительности испытываем, так это субъективные состояния мыслей и чувств, возникающие при объединении нейронных событий и названные философами квалиа . Но даже сами квалиа – один из видов эффекта нейронного связывания, объединения сигналов от бесчисленных нейронных сетей «низшего порядка». Все действительно сводится к электрохимическому процессу нейронного потенциала действия, или к возбуждению нейронов и установлению связи друг с другом с передачей информации. Как им это удается? Опять-таки благодаря химии. Связь между нейронами возникает в немыслимо крохотной синаптической щели между ними. Когда потенциал действия нейрона устремляется по аксону и достигает его терминалей, он вызывает выброс в синапс мельчайших порций химических трансмиттерных веществ (ХТВ). Полученные соединяющимися нейронами ХТВ действуют как ВПСП, меняя напряжение и проницаемость постсинаптического нейрона, тем самым вызывая его возбуждение и распространение его потенциала действия вниз по аксону до терминалей, где он выбрасывает свои ХТВ в следующий синаптический зазор, и так далее по всей линии нейронной сети. Когда мы ушибаем палец ноги, сигнал боли проходит от болевых рецепторов в тканях нашего пальца ноги весь путь вверх до мозга, который замечает боль и передает сигнал другим участкам мозга, посылающим дополнительные сигналы в сокращающиеся мышцы, чтобы мы отдернули ногу от злополучного препятствия. Все это происходит так быстро, что кажется почти мгновенным. Существует много видов ХТВ. К самым известным относятся катехоламины , в том числе допамин, норадреналин (норэпинефрин) и адреналин (эпинефрин) . ХТВ действуют на постсинаптический нейрон, как ключ на замок. Если ключ подошел и повернулся, нейрон срабатывает; в противном случае дверь остается запертой, а постсинаптический нейрон невозбужденным. После возникновения процесса возбуждения большинство неиспользованных ХТВ возвращается в пресинаптический нейрон, где-либо используется повторно, либо разрушается моноаминоксидазой (МАО) в процессе так называемого первого поглощения . Если в синаптическом зазоре присутствует слишком много ХТВ, тогда остаток всасывается в постсинаптический нейрон в процессе второго поглощения . Наркотики воздействуют на синапсы, выброс ХТВ и последующие процессы поглощения. Например, амфетамины ускоряют выброс ХТВ в синапсы, тем самым ускоряя процесс нейронной коммуникации, потому и называются speed («скорость»). Резерпин, который некогда был обычным назначением при психозах, разрушает пузырьки с ХТВ в пресинаптическом нейроне, поэтому МАО уничтожают их еще до использования, в итоге замедляют работу нейронных сетей, контролируют маниакальные состояния, гипертензию и другие симптомы гиперактивности нервной системы. Кокаин блокирует первое поглощение, поэтому ХТВ просто задерживаются в синапсе и способствуют ускоренному возбуждению нейронов, доводят нейронные сети до состояния взвинченности – вспомните Робина Уильямса с микрофоном перед аудиторией; в сущности, сам Уильямс в значительной мере приписывает успех своих комедий в 1980-х годах собственной кокаиновой зависимости. Как один из самых распространенных ХТВ, допамин играет решающую роль в беспрепятственной коммуникации между нейронами и мышцами, а когда его недостаточно, у пациентов наблюдаются потеря регуляции моторики и неудержимая дрожь. Эти проявления называются болезнью Паркинсона, один из методов лечения которой – L-dopa, агонист допамина, стимулирующий его выработку. Как нам построить всю систему снизу доверху, начиная с химических трансмиттерных веществ, таких как допамин, и связывая сигналы в единую систему убеждений? Посредством поведения. Напомню, что первичная функция мозга – управлять телом и помогать ему выжить. Один из способов сделать это – посредством ассоциативного обучения, или паттерничности. Это и есть связующее звено между нейронным потенциалом действия и человеческими поступками.
Допамин, наркотик веры
Из всех химических трансмиттерных веществ, плещущихся у нас в мозге, допамин, по-видимому, самым непосредственным образом связан с нейронными коррелятами веры. В сущности, допамин играет решающую роль в ассоциативном обучении и в подкрепляющей системе мозга, которую Скиннер открыл, применяя свой метод выработки условного рефлекса, когда любому подкрепленному поведению было свойственно повторяться. По определению, подкрепление – то, что служит наградой для организма, то есть побуждает мозг заставлять тело повторять это поведение, чтобы получить еще одно позитивное подкрепление. Вот как это происходит. В стволе разделенного головного мозга, одной из наиболее древних с точки зрения эволюции областей мозга, которая есть у всех позвоночных, имеются полости или карманы с приблизительно 15–24 тысячами вырабатывающих допамин нейронов с каждой стороны, длинные аксоны которых соединяются с другими областями мозга. Эти нейроны стимулируют выброс допамина всякий раз, когда полученное вознаграждение оказывается больше ожидаемого, в итоге индивид повторяет конкретное поведение. Выброс допамина – одна из форм предоставления информации, сообщение организму: «Сделай это еще раз». Допамин создает ощущение удовольствия, которым сопровождается решение задачи или достижение цели, в итоге организм хочет повторить то же самое поведение, будь то выжимание штанги, нажатие клавиши или манипуляции с рычагом механизма. Ты получаешь отклик (подкрепление), а твой мозг – дозу допамина. Поведение – Подкрепление – Поведение. Повторяющаяся последовательность . Однако у допаминовой системы есть свои плюсы и минусы. К плюсам можно отнести то, что допамин имеет отношение к пучку нейронов размером с орешек арахиса, расположенному посреди мозга и называющемуся nucleus accumbens (NAcc) – прилежащим ядром, которое, как известно, ассоциируется с наградой и удовольствием. В сущности, допамин, по-видимому, служит топливом этому так называемому центру удовольствия мозга, участвующему в «кайфе», который вызывают как кокаин, так и оргазм. «Центр удовольствия» был открыт в 1954 году Джеймсом Олдсом и Питером Милнером из Университета Макгилла, которые случайно вживили электрод в NAcc крысы и обнаружили, что грызун резко возбудился. Затем ученые сконструировали аппарат, который при нажатии крысой на планку создавал небольшую электрическую стимуляцию той же области мозга. Крысы давили на планку, пока не падали в изнеможении, даже забывали про пищу и воду.[104]Тот же эффект с тех пор был выявлен у всех участвовавших в экспериментах млекопитающих, в том числе и у людей, которые перенесли операцию на мозге и получили стимуляцию NAcc. Свои ощущения они описывали словом «оргазм ».[105]Вот это и есть типичный образец позитивного подкрепления! К сожалению, у допаминовой системы есть и минусы, а именно развивающаяся зависимость. Наркотики, вызывающие привыкание, играют роль сигнала награды, поступающего в допаминовые нейроны. Азартные игры, порнография, такие наркотики, как кокаин, способны вызвать в мозге ответный прилив допамина. Тот же эффект дают идеи, вызывающие зависимость, особенно неудачные идеи вроде тех, которые пропагандируют культы, например, призывающие к массовым самоубийствам (вспомните Джонстаун и «Небесные врата»), или религии, поощряющие действия террористов-смертников (вспомните теракты 11 сентября и 7 июля). Важное предостережение насчет допамина : нейробиологи делают четкое различие между «предпочтением» (удовольствием) и «желанием» (мотивацией), и в настоящее время продолжаются оживленные споры о том, чему именно способствует допамин – стимуляции удовольствия или мотивации поведения. Позитивное подкрепление может привести к повторам поведения, поскольку вызывает приятные ощущения (предпочтение, или чистое удовольствие от полученной награды) или неприятные ощущения, если поведение не повторяется (желание, или мотивация избегать беспокойства из-за неполучения награды). Первая награда связана с чистым удовольствием от, допустим, оргазма, вторая – с беспокойством, которое ощущает зависимый человек, когда получение следующей дозы внушает сомнения. Исследования, на которые я ссылаюсь выше, подтверждают предположение об удовольствии, однако по результатам новых исследований ученые склоняются к мотивации.[106]Нейробиолог Рассел Полдрак из Калифорнийского университета в Лос-Анджелесе рассказывал мне, что вновь полученные данные подразумевают «роль допамина скорее в мотивации, чем в удовольствии как таковом, в то время как опиоидная система, по всей видимости, играет центральную роль в удовольствии». Например, он указывает, что «можно блокировать допаминовую систему у крыс, и они все равно будут радоваться наградам, но не захотят стараться ради их получения».[107]Это трудноуловимое, но важное отличие, однако в целях нашего понимания нейронных коррелятов веры центральным является тот момент, что допамин подкрепляет поступки, убеждения и паттерничность и таким образом является одним из первичных «наркотиков веры». Связь между допамином и верой была установлена в ходе экспериментов, проведенных Питером Браггером и его коллегой Кристиной Моор в Бристольском университете, Англия. Исследуя нейрохимию суеверий, магического мышления и веры в паранормальные явления, Браггер и Моор обнаружили, что люди с высоким уровнем допамина с большей вероятностью находят смысл в совпадениях и усматривают значения и закономерности там, где их нет. Например, в одном исследовании сравнивали двадцать человек, объявивших, что они верят в призраков, богов, духов и заговоры, с двадцатью участниками, которые объявили о своем скептическом отношении к подобной вере. Всем участникам показали ряд слайдов с человеческими лицами, среди которых были как нормальные, так и «перепутанные», например, на некоторых глаза, уши или носы относились к другим лицам. В следующем эксперименте на экране вспыхивали существующие и произвольно составленные слова. В целом ученые обнаружили, что верующие с гораздо большей вероятностью, чем скептики, по ошибке принимали «перепутанное» лицо за настоящее, а придуманное слово – за обычное.
Допамин – наркотик веры. Люди с высоким уровнем допамина с большей вероятностью находят смысл в совпадениях и усматривают значения
и закономерности там, где их нет. Во второй части того же эксперимента Браггер и Моор дали всем сорока участникам L-dopa, препарат, который назначают пациентам с болезнью Паркинсона, чтобы повысить уровень допамина в мозге. После этого показ слайдов с лицами и словами повторили. Прилив допамина вызывал и у верующих, и у скептиков стремление воспринимать «перепутанные» лица, а также придуманные слова как обычные. Это свидетельствует о том, что паттерничность может ассоциироваться с высоким уровнем допамина в мозге. Любопытно, что на скептиков L-dopa действовал сильнее, чем на верующих. Иначе говоря, повышенный уровень допамина, по-видимому, уменьшал скептицизм скептиков эффективнее, чем усиливал веру верующих.[108]Почему? В голову приходят два возможных объяснения: (1) возможно, уровень допамина у верующих и без того выше, чем у скептиков, значит, последние острее ощущают его влияние; или (2) возможно, склонность верующих к паттерничности уже настолько высока, что эффект допамина у них ниже, чем у скептиков. Дополнительные исследования показали, что люди, заявляющие о своей вере в паранормальные явления, по сравнению со скептиками демонстрировали повышенную склонность усматривать «закономерности, или паттерны, в шумах»[109]и приписывать смысл произвольным связям, существующим по их мнению.[110]
Услышать в шуме сигнал
Так что же именно делает допамин, усиливая веру? Согласно одной теории, пропагандируемой Моор, Браггером и их коллегами, допамин повышает соотношение «сигнал-шум», то есть количество сигналов, который ваш мозг выявляет в фоновом шуме.[111]Такова проблема обнаружения ошибок, связанная с паттерничностью. Соотношение «сигнал-шум», в сущности, и есть проблема паттерничности – поиск значимых закономерностей как в исполненных смысла, так и в бессмысленных шумах. «Сигнал-шум» – соотношение паттернов, которые ваш мозг выявляет в фоновом шуме независимо от того, настоящие это паттерны или мнимые. Каким образом допамин влияет на этот процесс? Допамин усиливает способность нейронов передавать сигналы от одного к другому. Как? Выступая в роли агониста (в противоположность антагонисту), или вещества, усиливающего активность нейронов, допамин соединяется с особыми участками молекул рецепторов в синаптической щели нейронов, как ХТВ, обычно связывающиеся с ними.[112]При этом увеличивается уровень срабатывания нейронов в связи с распознанием паттерна, а это означает, что количество синаптических связей между нейронами скорее всего увеличится в ответ на воспринятый паттерн, тем самым впечатывая воспринятые паттерны в долгосрочную память благодаря реальному физическому росту новых нейронных соединений и усилению прежних синаптических связей. Прилив допамина вызывает усиление обнаружения паттернов; ученые выяснили, что агонисты допамина не только способствуют обучению, но и в больших дозах могут спровоцировать симптомы психоза, такие, как галлюцинации, возможно, связанные с тонкой гранью между креативностью (избирательная паттерничность) и безумием (неизбирательной паттерничностью). Все зависит от дозы. Если она слишком велика, скорее всего, возникнут ошибки первого типа, ложноположительное срабатывание, при которых мы видим связи там, где их на самом деле нет. Если доза слишком мала, возникают ошибки второго типа, ложноотрицательное срабатывание, при которых мы упускаем из виду реально существующие связи. Все дело в соотношении «сигнал-шум».
|
stydopedia.ru
Синаптогенез — Википедия
Синаптогенез — процесс формирования синапсов между нейронами в нервной системе. Синаптогенез происходит на протяжении всей жизни здорового человека, а взрыв формирования синапсов, т. н. избыточный синаптогенез (exuberant synaptogenesis), наблюдается на ранних стадиях развития головного мозга.[1] Синаптогенез особенно важен в ходе критического периода развития особи (в биологии развития, такого периода, когда нервная система особенно чувствительна к экзогенным стимулам), когда имеет место быть интенсивное протекание синаптического прунинга ввиду конкуренции нейронов и их синапсов за нейрональные факторы роста. Синапсы, которые не участвуют или участвуют слабо в обработке информации, как и неиспользуемые или малоиспользуемые отростки нейронов, не получат должного развития и будут отсеяны в дальнейшем.[2]
Формирование нервно-мышечных синапсов[править | править код]
Функция[править | править код]
Нервно-мышечное соединение (НМС) из всех типов синапсов наиболее хорошо изучено, оно представляет собой простую и доступную структуру для манипулирования и наблюдения. Этот синапс сам по себе состоит из трех клеток: мотонейрона, миоцита, и шванновской клетки (нейролеммоцита). В нормально функционирующем синапсе сигнал от мотонейрона вызывает деполяризацию мембраны миоцита, что происходит с помощью нейротрансмиттера ацетилхолина. Ацетилхолин высвобождается аксоном мотонейрона в синаптическую щель, где он достигает холинэргических рецепторов на сарколемме, мембране миоцита. Связывание ацетилхолина с рецепторами приводит к открытию ионных каналов, деполяризации мембраны и последующему каскаду химических реакций, результатом чего будет сокращение мышцы. Снаружи синапс инкапсулирует и изолирует миелиновая оболочка шванновской клетки.[3] Еще одним важным элементом центральной и периферической нервной системы являются астроциты. Хотя первоначально и полагалось, что их функция лишь структурная, сейчас известно, что они играют важную роль в функциональной пластичности синапсов.[4]
Происхождение и судьба клеток[править | править код]
В ходе развития каждый из трех типов вышеупомянутых клеток возникает из разных регионов развивающегося эмбриона. Отдельные миобласты развиваются из мезодермы и объединяются, формируя полиэнергидное (многоядерное) мышечное волокно. Во время или вскоре после образования волокна, мотонейроны нервной трубки начинают формировать первые контакты с ним. Шванновские клетки возникают из нервного гребня и направляются вместе с аксонами к месту назначения. По достижении мышечного волокна, они образуют свободное, немиелинизированное покрытие иннервирующих мышцы аксонов. Движение аксонов (а т. о. и клеток Шванна) управляется конусом роста, нитевидной проекцией аксона, которая активно ориентируется на нейротрофины, высвобождаемые мышечным волокном.
Исследование паттернов образования и распределения нервно-мышечных синапов показывает, что существенная часть соединений локализуется в средней точке мышечного волокна. Хотя может показаться, что аксоны изначально нацеливаются на этот локус, несколько факторов говорят об обратном. По-видимому, после первоначального контакта с аксоном, молодое волокно продолжает расти симметрично от точки иннервации. В сочетании с тем фактом, что высокая плотность холинэргических рецепторов является результатом контакта с аксоном, а не его причиной, паттерны формирования мышечных волокон можно связать как с индивидуальным миопатическим ростом, так и с иннервацией волокна аксонами.
Трансдукция сигнала по первичному контакту между мотонейроном и мышечным волокном начинается почти сразу, но передаваемый импульс очень слабый. Существуют свидетельства того, что Шванновские клетки, с помощью небольших сигнальных молекул, могут усиливать импульсы путем увеличения интенсивности спонтанного высвобождения нейромедиаторов.[5] Примерно через неделю мотонейрон и мышечное волокно проходят процесс дифференциации, чем завершается формирование полностью функционального синапса. Иннервация изначальным аксоном имеет решающие значение, поскольку последующие за ним аксоны имеют высокую склонность к формированию контактов с уже устоявшимся, стабилизированным синапсом.
Послесинаптическая дифференциация[править | править код]
Наиболее заметное изменение в мышечном волокне, возникающее после контакта с мотонейроном, — это повышенная концентрация холинэргических рецепторов на плазмалемме в месте контакта нейрона и миоцита. Это увеличение числа рецепторов обеспечивает более эффективную передачу синаптических сигналов, что в свою очередь ведет к формированию более развитого синапса. Плотность рецепторов составляет >10,000/мкм2 в центре и приблизительно 10/мкм2 на периферии. Такая высокая концентрация холинэргических рецепторов в синапсе достигается посредством их кластеризации, повышенной регуляции экспрессии соответствующих генов в ядрах пост-синаптических клеток и пониженной регуляции экспрессии этих генов в ядрах пре-синаптических клеток. Сигналами, которые инициируют послесинаптическую дифференциацию, могут быть нейротрансмиттеры, высвобождаемые непосредственно из аксона на мышечное волокно, или они могут возникать сторонним образом, попадая в синаптическую щель извне.[6]
Кластеризация[править | править код]
Кластеризация на пост-синаптической мембране происходит во многом благодаря протеогликану Агрину (Agrin), который экскретируется аксоном мотонейрона и способствует мультимеризации рецепторов. Агрин связывается с рецепторной мышечно-специфичной киназой (MuSK) на пост-синаптической мембране, а это в свою очередь приводит к последующей активации цитоплазматического белка Рапсина (Rapsyn). Именно рапсин содержит домены, позволяющие ему кластеризовать на мембране множество холинэргических рецепторов: мышечные клетки мутантных на этот белок мышей не были способны формировать кластеры холинэргических рецепторов.
Синапс-специфическая транскрипция[править | править код]
Повышенная концентрация холинэргических рецепторов достигается не только за счет манипуляций с уже существующими синаптическими компонентами. Аксон путём сигналинга способен на уровне процесса транскрипции осуществлять регуляцию экспрессии генов в ядре мышечной клетки. Всё это также происходит для того, чтобы повысить интенсивность локальной кластеризации рецепторов на мембране. В частности, в сигналинге задействованы две молекулы: белок семейства кальцитонинов (Calcitonin gene-related peptide) и представитель семейства белков-нейрегулинов (названных так за свою роль в регуляции нейрональных молекулярных процессов). Эти два белка вызывают каскад химических реакций с участием различных киназ, что в конечном итоге ведет к активации транскрипции генов холинэргических рецепторов.[7]
Внесинаптическая репрессия[править | править код]
Новообразованные синапсы способны генерировать электрические сигналы, таким образом воздействующие на окружающие внесинаптические клетки, что те существенно сокращают экспрессию генов холинэргических рецепторов. Точность импульса, посылаемого аксоном на пост-синаптическую мембрану, достигается именно за счет одновременного снижения концентрации рецепторов вне синапса, и повышения их количества внутри него. Поскольку синапс начинает получать входную информацию почти сразу после образования нервно-мышечного соединения, аксон быстро генерирует потенциал действия и высвобождает ацетилхолин. Деполяризация, вызванная связыванием ацетилхолина с его рецепторами, инициирует сокращение мышцы, и, одновременно с этим, ингибирование транскрипции рецепторных генов в клетках мышечного волокна вокруг синапса. Следует заметить, что эффект действует на расстоянии, а рецепторы, сидящие непосредственно на пост-синаптической мембране, репрессии не подвергаются.
Досинаптическая дифференциация[править | править код]
Хотя механизмы, регулирующие досинаптическую дифференциацию, неизвестны, изменения, происходящие в развивающейся аксональной терминали, охарактеризованы достаточно хорошо. Увеличивается объем и площадь синаптического контакта, объем везикул, концентрация везикул в активной зоне синапса, а также поляризация пресинаптической мембраны. Эти изменения, как считается, опосредованы нейротрофином и различными молекулами клеточной адгезии, высвобождающимися миоцитом; это подчеркивает важность связи между мотонейроном и мышечным волокном в процессе синаптогенеза. Считается, что именно изменение экспрессии генов и перераспределение составляющих компонентов синапса — ключевые моменты до- и послесинаптической дифференциации. В пользу этого также свидетельствует усиление экспрессии генов везикулярных белков вскоре после формирования синапса, а также их аклокализация в аксональной терминали.
Созревание синапса[править | править код]
Незрелые синапсы обнаруживают множественную иннервацию ввиду высокой склонности новых аксонов связываться с уже существующими синапсами. По мере созревания, синапсы обособляются и, в конечном счете, на входе остается только один аксон; это происходит в процессе, называемом элиминация синапса. Кроме того, путём инвагинации постсинаптическая сторона увеличивает площадь принимающей нейротрансмиттеры поверхности. На ранних этапах онтогенеза, шванновские клетки формируют рыхлую, немиелинизирующую оболочку, покрывающую группу синапсов, однако по мере роста и развития, пределы компетенции отдельно взятого нейролеммоцита сужаются до одного синапса.
Элиминация синапса[править | править код]
Элиминация синапса, как один из механизмов синаптического прунинга, предполагает под собой процесс, напрямую зависящий от активности конкурирующих между собой аксонов. В случае конкретно взятого синапса, если частота импульсов на входе достаточна для создания потенциала действия, то это будет провоцировать экспрессию генов белков-синаптотрофинов в ядре пост-синаптического миоцита, и эти белки будут поддерживать данный синапс. Возникает механизм положительной обратной связи, недоступный менее востребованным синапсам, вследствие этого обреченным на элиминацию. Также было высказано предположение, что в дополнение к синаптотропинам, высвобождаемым в активных синапсах, деполяризация их постсинаптической мембраны вызывает высвобождение синаптотоксинов, ингибирующих аксоны в неактивных синапсах.
Специфика формирования синапсов[править | править код]
Примечательным аспектом синаптогенеза является тот факт, что мотонейроны способны различать быстрые и медленные мышечные волокна; быстрые мышечные волокна иннервируются «быстрыми» мотонейронами, а мышечные волокна с медленным сокращением иннервируются «медленными» мотонейронами. Существуют два гипотетических пути, согласно которым аксоны мотонейронов достигают этой специфичности; эти пути отличаются между собой в ключевом моменте — степени избирательности, в которой аксон и иннервируемое им мышечное волокно взаимодействуют друг с другом, степени обработки аксоном входных данных от миоцита. Первая гипотеза предполагает, что аксоны выбирают «быстрый» или «медленный» путь в зависимости от сигнальных веществ, различных у быстрых и медленных мышечных волокон. Кроме того, селективность наблюдается еще на ранних стадиях, когда аксоны ориентируются по отношению к мышечным волокнам так, чтобы конкретный аксон иннервировал конкретную мышечную клетку. Вторая гипотеза заключается в том, что движение аксона и его связь с конкретным миоцитом обеспечиваются внеклеточным матриксом, омывающим аксон, то есть не конечная цель аксона, но окружающая среда определяет его судьбу. Наконец, может быть и такое, что формирование нервно-мышечного соединения не предопределено заранее, а мышечные волокна приобретают свои быстрые или медленные характеристики уже после, в зависимости от характеристик иннервирующего аксона. Если так, то «быстрые» мотонейроны способны трансформировать всякое мышечное волокно в быстро-сокращающееся. Существует множество свидетельств в пользу всех описанных возможностей, что заставляет склоняться к мысли об их комбинации в процессе формирования синапса.
Данные, полученные в исследованиях нервно-мышечных синапсов, можно попытаться экстраполировать на процессы синаптогенеза в ЦНС, даже несмотря на то, что исследования последних имеют намного более поздний характер. Эти два типа соединений имеют много общего. Оба типа синапса имеют консервативное базовое строение: пре-синаптическая нервная терминаль, синаптическая щель и принимающая постсинаптическая клетка. Оба типа синапса содержат высокую концентрацию везикул в активном центре, кластеры рецепторов на принимающей мембране, и, наконец, оба они изолированы специальными глиальными клетками. Процессы, происходящие в них, также достаточно консервативны. Это и кластеризация рецепторов, и локализованное повышение интенсивности синтеза белка в активных центрах, и синаптический прунинг путем элиминации малоактивных синапсов.
Несмотря на вышеобозначенные сходства, существует принципиальная разница между этими двумя соединениями. Поскольку постсинаптической клеткой уже не выступает миоцит, в корне отличаются действующие рецепторы и их нейротрансмиттеры. Что еще более важно, нейроны в пределах ЦНС иннервированы многими другими нейронами одновременно, и должны уметь интегрировать и обрабатывать большое количество информации на входе, когда как мышечные волокна иннервируются одним нейроном и сокращаются в ответ на одиночный импульс. В сочетании с пластичностью, характерной для нейронных связей, нейронная сеть в ЦНС имеет огромный потенциал для усложнения.
Факторы, регулирующие синаптогенез в ЦНС[править | править код]
Сигналинг[править | править код]
Гомологом ацетилхолина и его рецепторов в ЦНС является глутамат и N-метил-D-аспартат-рецепторы (NMDA). Было показано, что активация NMDA-рецепторов инициирует каскад химических реакций, ведущих к формированию и стабилизации синапса. Повышенный уровень активности NMDA-рецепторов во время развития синапса позволяет увеличить приток кальция, который выступает в качестве вторичного сигнала. Помимо этого, факторами транскрипции активируются гены первичного ответа (ГПО), играющие важную роль в таких процессах, как, например, долговременная потенциация.[8] Функция рецепторов NMDA также связана с эстрогеновыми рецепторами в гиппокампе. Эксперименты, проведенные с эстрадиолом, показывают, что воздействие эстрогена значительно увеличивает плотность синапсов и концентрацию белка.[9]
Передача нервного импульса во время синаптогенеза зависит не только от активности отдельно взятого синапса, но и от окружающей этот синапс среды. Например, нейротрофический фактор головного мозга (BDNF) — продуцируется головным мозгом и регулирует несколько функций в развивающемся синапсе, включая усиление высвобождения нейротрансмиттеров, повышение концентрации везикул и биосинтез холестерина. Холестерин является важной структурной составляющей липидного бислоя, выступающим как место действия самых разных сигнальных взаимодействий. Мутантные на наличие BDNF организмы обнаруживают значительные дефекты в росте нейронов и формировании синапсов.[10] Помимо нейротрофинов, важную роль в процессе синаптогенеза играют также молекулы клеточной адгезии. Во многом синаптогенез зависит от корректного локального связывания одной адгезионной молекулы с другой. Показано, что дефекты в генах, кодирующих белки-нейролигины, одни из важнейших молекул клеточной адгезии, играющие роль в процессе стабилизации синапса, связаны с некоторыми случаями аутизма и умственной отсталости.[11] Наконец, многие из этих сигнальных процессов могут регулироваться с помощью матриксных металлопротеиназ (ММП).
Морфология[править | править код]
Высокодинамичными сайтами, принимающими одновременно несколько входящих импульсов, являются дендритные шипики. Этот морфологический динамизм обусловлен специфической регуляцией актинового цитоскелета, что, в свою очередь, позволяет регулировать образование синапсов.[12] Морфологически, дендритные шипики можно разделить на три основных вида: филоподии, тонкие шипики и грибовидные шипики. Филоподии инициируют контакт с аксонами других нейронов. Филоподии новообразованных нейронов имеют тенденцию к образованию контактов с аксонами, иннервирующими множество синапсов, когда как филоподии более старых нейронов связываются преимущественно с аксонами, иннервирующими одиночный синапс. Благодаря высокой динамичности, одни типы шипиков способны переходить в другие, например, филоподии легко трансформируются в грибоподобные шипики, которые являются первичными сайтами глутаматных рецепторов.[13]
Экологическое обогащение — термин, обозначающий стимуляцию развивающегося мозга физическим и социальным окружением. Экологическое обогащение приводит к тому, что эффективность процессов синаптогенеза существенно увеличивается. Так, выращенные в среде с таким эффектом крысы показывают увеличение количества синапсов на 25% в сравнении с контролем.[14][15] Этот эффект наблюдается независимо от того, возникает ли стимулирующая среда сразу после рождения,[16] вскоре после отлучения,[17] или во время взросления.[18] Не только для пирамидальных, но и для звездчатых нейронов экологическое обогащение вызывает всплеск синаптогенеза.[19]
В белковом семействе Wnt существует несколько эмбриональных морфогенов (веществ, определяющих морфогенез тканей), которые способствуют раннему формированию структур развивающегося эмбриона. Недавно появившиеся данные показывают вклад морфогенов этого семейства в синаптогенез и пластичность, причем семейство вносит вклад в синаптогенез как синапсов ЦНС, так и нервно-мышечных синапсов.
Центральная нервная система[править | править код]
В синапсах мозжечка Wnt-семейство индуцирует созревание пре- и постсинаптических окончаний. Эта область мозга содержит три основных типа нейронов: клетки Пуркинье, гранулярные клетки и клетки мшистых волокон. У клеток Пуркинье Wnt-3 стимулирует рост как дендритов, так и аксонов.[20][21] Гранулярные клетки экспрессируют Wnt-7а, который индуцирует рост и ветвление аксонов в клетках мшистых волокон. Ретроградная секреция Wnt-7a, направленная на мшистые волокна,вызывает увеличение конуса роста аксонов путем увеличения числа микротрубочек. Кроме того, ретроградный Wnt-7a-сигналинг вызывает накопление синаптических пузырьков и пресинаптических белков в активной зоне синапса. Внт-5А выполняет аналогичную функцию у постсинаптических гранулярных клеток; Wnt-5-сигналинг стимулирует сборку рецепторов на мембране, а также кластеризацию на мембране каркасного белка PSD-95.
В гиппокампе белки семейства Wnt, в сочетании с электрической активностью клеток, также способствует формированию синапсов. Wnt7b экспрессируется в созревающих дендритах, помимо этого, в формирующихся синапсах возрастает экспрессия рецепторов семейства Frizzled (Fz), специфических для Wnt-белков. Глутаматные NMDA-рецепторы активируются в ответ на экспрессию белков Wnt-2-типа. Долговременная потенциация вследствие активации NMDA-рецепторов и последующая Wnt-экспрессия ведет к кластеризации рецепторов Fz-5 типа на мембране пост-синаптических клеток. Кроме того, Wnt-7a и Wnt-2 типы сигналинга после опосредованной таким образом долговременной потенциации увеличивают интенсивность процесса ветвления дендритов, а также регулируют синаптическую пластичность. Блокирование экспрессии белков Wnt-семейства закономерно приводит к ингибированию этих процессов.[22]
Нервно-мышечный синапс[править | править код]
Подобные механизмы, опосредованные Wnt-семейством в ЦНС, хорошо прослеживаются и в нервно-мышечных соединениях. У Дрозофил мутации Wnt-5-рецептора в нервно-мышечных синапсах приводят к снижению количества и плотности активных центров. Основным нейротрансмиттером в этой системе является глутамат. Белки Wnt-семейства необходимы для правильной локализации глутаматергических рецепторов на постсинаптической мембране миоцитов. Мутации в белках этого семейства приводят к снижению интенсивности волны деполяризации после передачи импульса с аксона на мышечное волокно.
У позвоночных в нервно-мышечных синапсах экспрессия белка Wnt-11r способствует кластеризации холинэргических рецепторов на постсинаптической мембране мышечных клеток. Белок Wnt-3 экспрессируется мышечными волокнами и секретируется ретроградно на моторные нейроны. В моторных нейронах, Wnt-3 работает в паре с Агрином для стимуляции расширения конуса роста, ветвления аксональных терминалей и кластеризации синаптических пузырьков.
- ↑ Huttenlocher, P. R.; Dabholkar, A. S. Regional differences in synaptogenesis in human cerebral cortex (англ.) // The Journal of Comparative Neurology (англ.)русск. : journal. — 1997. — Vol. 387, no. 2. — P. 167—178. — DOI:10.1002/(SICI)1096-9861(19971020)387:2<167::AID-CNE1>3.0.CO;2-Z. — PMID 9336221.
- ↑ Comery T. A., Harris J. B., Willems P. J., etal. Abnormal dendritic spines in fragile X knockout mice: maturation and pruning deficits (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1997. — May (vol. 94, no. 10). — P. 5401—5404. — DOI:10.1073/pnas.94.10.5401. — PMID 9144249.
- ↑ Sanes J. R., Lichtman J. W. Development of the vertebrate neuromuscular junction (англ.) // Annu. Rev. Neurosci. : journal. — 1999. — Vol. 22. — P. 389—442. — DOI:10.1146/annurev.neuro.22.1.389. — PMID 10202544.
- ↑ Ullian EM, Christopherson KS, Barres BA. 2004. Role for glia in synaptogenesis. Glia 47(3):209-16.
- ↑ Cao G., Ko C. P. Schwann cell-derived factors modulate synaptic activities at developing neuromuscular synapses (англ.) // J. Neurosci. (англ.)русск. : journal. — 2007. — June (vol. 27, no. 25). — P. 6712—6722. — DOI:10.1523/JNEUROSCI.1329-07.2007. — PMID 17581958.
- ↑ Matrix metalloproteinases in brain development and remodeling: synaptic functions and targets (англ.) // J. Neurosci. Res. (англ.)русск. : journal. — 2007. — October (vol. 85, no. 13). — P. 2813—2823. — DOI:10.1002/jnr.21273. — PMID 17387691.
- ↑ Hippenmeyer S., Huber R. M., Ladle D. R., Murphy K., Arber S. ETS transcription factor Erm controls subsynaptic gene expression in skeletal muscles (англ.) // Neuron (англ.)русск. : journal. — Cell Press (англ.)русск., 2007. — September (vol. 55, no. 5). — P. 726—740. — DOI:10.1016/j.neuron.2007.07.028. — PMID 17785180.
- ↑ Ghiani C. A., Beltran-Parrazal L., Sforza D. M., etal. Genetic program of neuronal differentiation and growth induced by specific activation of NMDA receptors (англ.) // Neurochem. Res. (англ.)русск. : journal. — 2007. — February (vol. 32, no. 2). — P. 363—376. — DOI:10.1007/s11064-006-9213-9. — PMID 17191130.
- ↑ Jelks K. B., Wylie R., Floyd C. L., McAllister A. K., Wise P. Estradiol targets synaptic proteins to induce glutamatergic synapse formation in cultured hippocampal neurons: critical role of estrogen receptor-alpha (англ.) // J. Neurosci. (англ.)русск. : journal. — 2007. — June (vol. 27, no. 26). — P. 6903—6913. — DOI:10.1523/JNEUROSCI.0909-07.2007. — PMID 17596438.
- ↑ Suzuki S., Kiyosue K., Hazama S., etal. Brain-derived neurotrophic factor regulates cholesterol metabolism for synapse development (англ.) // J. Neurosci. (англ.)русск. : journal. — 2007. — June (vol. 27, no. 24). — P. 6417—6427. — DOI:10.1523/JNEUROSCI.0690-07.2007. — PMID 17567802.
- ↑ Zeng X., Sun M., Liu L., Chen F., Wei L., Xie W. Neurexin-1 is required for synapse formation and larvae associative learning in Drosophila (англ.) // FEBS Lett. (англ.)русск. : journal. — 2007. — May (vol. 581, no. 13). — P. 2509—2516. — DOI:10.1016/j.febslet.2007.04.068. — PMID 17498701.
- ↑ Proepper C., Johannsen S., Liebau S., etal. Abelson interacting protein 1 (Abi-1) is essential for dendrite morphogenesis and synapse formation (англ.) // EMBO J. (англ.)русск. : journal. — 2007. — March (vol. 26, no. 5). — P. 1397—1409. — DOI:10.1038/sj.emboj.7601569. — PMID 17304222.
- ↑ Toni N., Teng E. M., Bushong E. A., etal. Synapse formation on neurons born in the adult hippocampus (англ.) // Nat. Neurosci. : journal. — 2007. — June (vol. 10, no. 6). — P. 727—734. — DOI:10.1038/nn1908. — PMID 17486101.
- ↑ Diamond M. C., Krech D., Rosenzweig M. R. The Effects of an Enriched Environment on the Histology of the Rat Cerebral Cortex (англ.) // J. Comp. Neurol. (англ.)русск. : journal. — 1964. — August (vol. 123). — P. 111—120. — DOI:10.1002/cne.901230110. — PMID 14199261.
- ↑ Diamond M. C., Law F., Rhodes H., etal. Increases in cortical depth and glia numbers in rats subjected to enriched environment (англ.) // J. Comp. Neurol. (англ.)русск. : journal. — 1966. — September (vol. 128, no. 1). — P. 117—126. — DOI:10.1002/cne.901280110. — PMID 4165855.
- ↑ Schapiro S., Vukovich K. R. Early experience effects upon cortical dendrites: a proposed model for development (англ.) // Science. — 1970. — January (vol. 167, no. 3916). — P. 292—294. — DOI:10.1126/science.167.3916.292. — PMID 4188192.
- ↑ Bennett E. L., Diamond M. C., Krech D., Rosenzweig M. R. Chemical and Anatomical Plasticity Brain (англ.) // Science. — 1964. — October (vol. 146, no. 3644). — P. 610—619. — DOI:10.1126/science.146.3644.610. — PMID 14191699.
- ↑ Briones T. L., Klintsova A. Y., Greenough W. T. Stability of synaptic plasticity in the adult rat visual cortex induced by complex environment exposure (англ.) // Brain Res. (англ.)русск. : journal. — 2004. — August (vol. 1018, no. 1). — P. 130—135. — DOI:10.1016/j.brainres.2004.06.001. — PMID 15262214.
- ↑ Greenough W. T., Volkmar F. R. Pattern of dendritic branching in occipital cortex of rats reared in complex environments (англ.) // Exp. Neurol. (англ.)русск. : journal. — 1973. — August (vol. 40, no. 2). — P. 491—504. — DOI:10.1016/0014-4886(73)90090-3. — PMID 4730268.
ru.wikipedia.org