энтеробактерии, стафилококки, нефер-ментирующие бактерии) с определением чувствительности к бак-териофагам
Код услуги
77010200Описание
Кал на условно-патогенную микрофлору. Цель исследования.изучение флоры толстой кишки , обнаружение в кале условно — патогенной микрофлоры : энтеробактерий (кишечной палочки с нормальной и измененной ферментативной активностью, клебсиелл, энтеробактер, цитробактер, протеев, морганелл, гафний, сераций), стафилококков, неферментирующей флоры, синегнойной палочки. Определение чувствительности выделенной условно-патогенной микрофлоры к бактериофагам. Исследуемый материал.Кал. Показания к исследованю. Длительно протекающие кишечные расстройства. Затянувшийся период реконвалесценции после ОКИ. Дисфункция кишечнника у лиц , длительноо подвергающихся воздействию вредных факторов ( излучение, химиотерапия, гормональная терапия). Наличие системной воспалительной реакции или бактериемии. Частые респираторные инфекции. Аллергические заболевания.

Подготовка
Кал для исследования отбирается из средней порции в количестве 1-2 г (2 ложечки) в специальный стерильный контейнер. При запорах можно использовать слабительные средства только растительного происхождения (сена,слабительный чай,чернослив,свекла).Кал должен быть нормальной консистенции.Нельзя использовать клизму.Доставка в течение 2 часов, кроме воскресенья.Медицинский центр — Асклепий Бактериологические исследования
Исследуемый материал: Кал, ректальный соскоб
Сроки исполнения: 7 р.д.
Подготовка к исследованию:
Исследование рекомендуется проводить до начала приема антибиотиков и других антибактериальных химиотерапевтических препаратов.
Исключить прием слабительных препаратов, введение ректальных свечей, масел, ограничить (по согласованию с врачом) прием медикаментов, влияющих на перистальтику кишечника (белладонна, пилокарпин и др.
Об исследовании:
Микробиологическое исследование, позволяющее выявить в кале патогенную флору: возбудителей бактериальной дизентерии (шигеллеза), сальмонеллезов, в том числе брюшного тифа и паратифа, а также патогенные группы кишечной палочки (E. coli) – возбудителей эшерихиозов. Патогенные микроорганизмы не входят в состав нормальной микрофлоры и в норме у человека отсутствуют. При попадании в организм они, как правило, приводят к инфекционным заболеваниям.
Бактерии дизентерийной группы вызывают дизентерию (шигеллез) – острое инфекционное заболевание с фекально-оральным механизмом передачи возбудителя. Она сопровождается преимущественным поражением толстой кишки с диареей, лихорадкой и болями в животе. В стуле пациента зачастую обнаруживается кровь, слизь или гной.
К бактериям тифопаратифозной группы относятся Salmonella typhi (возбудитель брюшного тифа), Salmonella paratyphi A, B, C (возбудитель паратифа A, B, C соответственно), а также возбудители других сальмонеллезов.
Сдать анализ кала на дисбактериоз кишечника в Санкт-Петербурге
Краткое описаниеМикробиологическое исследование, которое позволяет оценить состав микрофлоры кишечника – концентрацию и соотношение «полезных», условно-патогенных и патогенных микроорганизмов в кале, определить специфическую активность основных пробиотиков в отношении выявленных микроорганизмов и их чувствительность к бактериофагам и антибиотикам.
 К нормальной микрофлоре кишечника («полезные» бактерии) относятся лактобактерии, бифидобактерии, энтерококки, кишечные палочки (типичные), анаэробная флора (бактероиды). Условно-патогенная флора – это энтеробактерии, неферментирующие бактерии, стафилококки, анаэробные бактерии (клостридии), грибы. Патогенными микроорганизмами являются сальмонеллы, шигеллы, патогенные эшерихии. При выявлении патогенных и/или условно-патогенных микроорганизмов будет определена их чувствительность к антимикробным препаратам (антибиотикам и бактериофагам) и пробиотикам. При обнаружении микроорганизмов, составляющих нормальную микрофлору, чувствительность к антибиотикам и бактериофагам не определяется, т.к. не имеет диагностического значения.
К нормальной микрофлоре кишечника («полезные» бактерии) относятся лактобактерии, бифидобактерии, энтерококки, кишечные палочки (типичные), анаэробная флора (бактероиды). Условно-патогенная флора – это энтеробактерии, неферментирующие бактерии, стафилококки, анаэробные бактерии (клостридии), грибы. Патогенными микроорганизмами являются сальмонеллы, шигеллы, патогенные эшерихии. При выявлении патогенных и/или условно-патогенных микроорганизмов будет определена их чувствительность к антимикробным препаратам (антибиотикам и бактериофагам) и пробиотикам. При обнаружении микроорганизмов, составляющих нормальную микрофлору, чувствительность к антибиотикам и бактериофагам не определяется, т.к. не имеет диагностического значения.Срок выполнения от 5 до 7 суток
Синонимы (rus) Антагонистическая активность пробиотиков, Определение пробиотиков-антагонистов, Определение чувствительности к бактериофагам и антибиотикам
Методы Микробиологический метод
Подготовка к исследованию
- Исследование рекомендуется проводить до начала приема антибиотиков и других антибактериальных химиотерапевтических препаратов.


Тип биоматериала
Кал
Сдать анализ кала на дисбактериоз кишечника
Нормальная кишечная микрофлора включает в себя более 500 различных видов микроорганизмов, биологическая роль которых заключается в переваривании пищи, регулировании иммунитета, усвоении микроэлементов, синтезе витаминов, участии в различных видах обмена, стимулировании кишечной перистальтики.
Выделяют обязательную “полезную” микрофлору (лактобактерии, бифидобактерии, энтерококки, некоторые кишечные палочки), условно-патогенную флору (к ней относятся, например, клостридии, стафилококки, грибы), а также абсолютно патогенную флору (сальмонеллы, шигеллы и др.
Дисбактериоз проявляется нарушением общего состояния (интоксикация, обезвоживание), снижением массы тела, симптомами поражения слизистых оболочек желудочно-кишечного тракта, расстройством работы кишечника, нарушением пищеварения в различных отделах кишечника, снижением иммунитета. Такие выраженные симптомы, как со стороны ЖКТ, так и со стороны общей реакции организма, объясняются крайне важным влиянием микрофлоры кишечника на все системы и органы.
- Длительный прием антибактериальных препаратов, НПВС, а также средств, подавляющих иммунитет (например, в лечении онкологических заболеваний, системных болезней)
- Длительный прием алкоголя и сильнодействующих веществ
- Наличие заболеваний ЖКТ, например, неспецифический язвенный колит, синдром мальабсорбции
Это микробиологическое исследование кала, которое дает возможность обнаружить качественные и количественные нарушения состава кишечной микрофлоры, а также патогенную микрофлору, вызывающую различные заболевания. Выявляемые микроорганизмы: бифидобактерии, лактобациллы, стафилококки, стрептококки, энтеробактерии, энтерококки, клебсиеллы, дрожжеподобные грибы и др. Исследование является высокочувствительным и точно отражает состав кала — оценивается количество облигатных бактерий, условно-патогенных, абсолютных патогенов, если они имеются. Анализ на дисбактериоз кишечника (микробиологическое исследование кала) позволяет оценить чувствительность выявленных микроорганизмов к бактериофагам. Определение чувствительности проводится при наличии чрезмерного роста условно-патогенной флоры и абсолютных патогенов. Если же микрофлора представлена полезной флорой и нормальным количество условно-патогенных микроорганизмов, то чувствительность к бактериофагам не проверяют, так как данное состояние является вариантом нормы и лечение не требуется. Но в случаях, когда имеет место дисбактериоз или же инфекция, крайне важно выявить, к каким бактериофагам чувствительны те или иные патогены. Следует учитывать особенность состава микрофлоры, определяющейся в кале, и микрофлоры, которая обитает непосредственно на слизистой кишечника — существуют некоторые отличия, но в целом анализ на дисбактериоз отражает состояние биоптата ЖКТ.
Выявляемые микроорганизмы: бифидобактерии, лактобациллы, стафилококки, стрептококки, энтеробактерии, энтерококки, клебсиеллы, дрожжеподобные грибы и др. Исследование является высокочувствительным и точно отражает состав кала — оценивается количество облигатных бактерий, условно-патогенных, абсолютных патогенов, если они имеются. Анализ на дисбактериоз кишечника (микробиологическое исследование кала) позволяет оценить чувствительность выявленных микроорганизмов к бактериофагам. Определение чувствительности проводится при наличии чрезмерного роста условно-патогенной флоры и абсолютных патогенов. Если же микрофлора представлена полезной флорой и нормальным количество условно-патогенных микроорганизмов, то чувствительность к бактериофагам не проверяют, так как данное состояние является вариантом нормы и лечение не требуется. Но в случаях, когда имеет место дисбактериоз или же инфекция, крайне важно выявить, к каким бактериофагам чувствительны те или иные патогены. Следует учитывать особенность состава микрофлоры, определяющейся в кале, и микрофлоры, которая обитает непосредственно на слизистой кишечника — существуют некоторые отличия, но в целом анализ на дисбактериоз отражает состояние биоптата ЖКТ.
ᐈ Диагностика и лечение дисбактериоза кишечника в Санкт-Петербурге
Дисбактериоз кишечника – очень распространенная и при этом недостаточно оцениваемая в современном обществе проблема. Дисбаланс кишечной флоры отмечается почти у 90% людей, но в большинстве случаев он протекает малосимптомно и остается не диагностированным. А это негативно сказывается на качестве пищеварения и становится предпосылкой для развития и рецидивирования нарушений и заболеваний. Поэтому выявление и лечение дисбактериоза кишечника – это путь к общему оздоровлению организма.
Дисбактериоз – это болезнь или вариант нормы?
Дисбактериозом (дисбиозом) называют стойкие нарушения кишечной микрофлоры, со значительным изменением баланса между количеством и активностью условно-патогенных и полезных для человека микроорганизмов. В результате просвет кишечника становится местом обитания большого количества бактерий и грибов, которые нарушают работу желудочно-кишечного тракта и способны негативно влиять на состояние всего организма.
Следует понимать, что речь идет не о воздействии конкретного болезнетворного микроорганизма (как при кишечных инфекциях). Дисбактериоз – это системное нарушение нормального состояния кишечника. Негативное влияние оказывает и увеличение количества условно-патогенных микроорганизмов, и недостаток полезных бактерий-помощников.
Человеческий организм в норме не является стерильным. На поверхности нашей кожи, в кишечнике и в большинстве других полых органов обитает большое количество микроорганизмов. Некоторые из них нейтральны для человека, другие при определенных условиях и снижении иммунитета способны проявлять агрессию. Такая флора называется условно-патогенной, в норме она малоактивна и не представляет опасности. Лечиться для полного избавления от нее не надо.
В просвете кишечника обитают и полезные для нашего организма бактерии: лактобактерии, бифидобактерии, энтеробактерии и др. Они выполняют много функций:
- участвуют в окончательном переваривании пищи;
- синтезируют некоторые витамины;
- поддерживают необходимую кислотность в просвете кишечника;
- влияют на моторику пищеварительного тракта;
- регулируют обмен холестерина;
- способствуют образованию необходимых антител;
- контролируют количество и активность условно-патогенной флоры, подавляя рост гноеродных и гнилостных бактерий.
Уже развившийся дисбактериоз толстой кишки или тонкого кишечника иногда постепенно самостоятельно нейтрализуется, но чаще всего он принимает затяжное течение. Такое состояние не является физиологическим (естественным) для человека. Это требующая коррекции патология, хотя ее и не приравнивают к самостоятельным заболеваниям.
Почему такое развивается
У взрослых дисбактериоз всегда является вторичным, речь идет о нарушении баланса уже существующей в кишечнике микрофлоры. А вот у детей первых месяцев жизни желудочно-кишечный тракт еще только заселяется микроорганизмами, этот процесс может происходить неравномерно, с временным преобладанием условно-патогенных бактерий и грибов. Такой дисбактериоз у грудного ребенка называют первичным.
Полезная микрофлора кишечника чувствительна к разнообразным поступающим извне веществам, к ферментному балансу в пищеварительном тракте и к составу пищи. Поэтому причин для развития дисбактериоза достаточно много:
- Прием антибактериальных средств, кишечных антисептиков, препаратов с широким антимикробным действием. Причем значение имеют и назначенные врачом лечебные курсы, и бессистемное самолечение.
- Несбалансированное питание, с малым количеством клетчатки и сложных углеводов, с избытком жареного жирного красного мяса и простых углеводов.
- Злоупотребление слабительными средствами и клизмами. Часто повторяющиеся кишечные инфекции, синдром раздраженного кишечника, хроническая диарея другого происхождения
- Частый прием сорбентов.
- Хронические заболевания кишечника и других органов пищеварения.
- Эндокринные заболевания.
- Лучевая терапия, химиотерапия, некоторые другие методы лечения с цитостатическим эффектом.
 Причем значение имеют и назначенные врачом лечебные курсы, и бессистемное самолечение.
Причем значение имеют и назначенные врачом лечебные курсы, и бессистемное самолечение.
Усугубляют ситуацию хронические стрессы, недосыпание, гиподинамия, вредные привычки и многие другие факторы. Они действуют опосредованно, ухудшая работу кишечника и иммунной системы, что в итоге негативно сказывается на составе кишечной флоры и ухудшает саморегуляцию. Грамотное лечение дисбактериоза должно учитывать все эти моменты.
Основные проявления
Дисбаланс кишечной микрофлоры в первую очередь отражается на качестве пищеварения, поэтому самое частое проявление заболевания – это кишечное расстройство. Но дисбактериоз приводит и к другим нарушениям, что, к сожалению, не всегда принимается во внимание при подборе схемы лечения.
Самые частые симптомы и признаки дисбактериоза:
- Нарушения работы ЖКТ. Людей с дисбиозом кишечника могут беспокоить урчание и периодический дискомфорт в животе, метеоризм, отрыжка, кишечные расстройства (запоры, поносы или их чередование). Такие признаки могут иметь различную интенсивность, возникать периодически или беспокоить основную часть времени. Наиболее яркую и дискомфортную симптоматику обычно дает дисбактериоз тонкой кишки.
- Неприятный запах изо рта, необычный привкус.
- Признаки гиповитаминоза, а иногда и анемии. Это связано с нарушением процесса переваривания и усвоения питательных веществ, со снижением синтеза витаминов группы В полезной кишечной микрофлорой, со повторяющейся диареей (поносом).
- Ухудшение состояния кожи и волос, склонность к дерматитам.

Анализ на дисбактериоз желательно также сдавать людям с частыми аллергическими реакциями, аутоиммунными заболеваниями, атопическим дерматитом, резистентной (устойчивой к лечению) бронхиальной астмой и псориазом. Дело в том, что дисбаланс кишечной флоры меняет реактивность иммунной системы, что может поддерживать симптомы заболеваний с аллергической и смешанной природой.
Как выявить дисбактериоз
План комплексного обследования при дисбиозе кишечника включает:
-
Микробиологический анализ кала. Это базовое исследование, позволяющее оценить состав кишечной микрофлоры. По назначению врача одновременно определяется чувствительность микроорганизмов к противомикробным препаратам и бактериофагам.
-
Копрологическое исследование для оценки качества переваривания пищи.
-
УЗИ органов брюшной полости.
Относится к вспомогательным методам, используется для выявления структурных изменений органов ЖКТ.
-
Общеклиническая лабораторная диагностика (общие анализы крови и мочи, биохимический анализ крови). Позволяет определить наличие и выраженность воспалительных реакций, выявить анемию, оценить функциональное состояние печени, поджелудочной железы, почек.
-
Эндоскопическое исследование: ФГДС, ФКС. Позволяет осмотреть поверхность слизистой оболочки органов пищеварения, взять биопсию для гистологического исследования измененных тканей, провести исследование причин дисбактериоза. Использование эндоскопии значительно повышает качество диагностики заболеваний ЖКТ.
 Относится к вспомогательным методам, используется для выявления структурных изменений органов ЖКТ.
Относится к вспомогательным методам, используется для выявления структурных изменений органов ЖКТ.
Существуют также лабораторные тесты для выявления патологических метаболитов и веществ, выделяемых патогенными бактериями. Но в повседневной клинической практике они используются редко.
Принципы лечения
Грамотно подобранная терапия дисбактериоза направлена не только на коррекцию основных симптомов и выявленных нарушений. Первостепенная задача лечения – восстановление физиологического баланса микрофлоры. Для этого необходимо подавить активность патогенных бактерий и грибов, заселить ЖКТ полезными бактериями и создать условия для их размножения.
Первостепенная задача лечения – восстановление физиологического баланса микрофлоры. Для этого необходимо подавить активность патогенных бактерий и грибов, заселить ЖКТ полезными бактериями и создать условия для их размножения.
Лечение дисбактериоза подбирается индивидуально и обычно включает:
- Соблюдение диеты для улучшения процесса переваривания, обеспечения организма необходимыми веществами.
- Препараты для подавления патогенной флоры, с учетом чувствительности к ним выделенных микроорганизмов. Это могут быть антибиотики, противомикробные средства, бактериофаги.
- Эубиотики – средства, которые содержат специально выведенные штаммы жизнеспособных полезных бактерий и вещества для их питания и роста.
- Энтеросорбенты, осаждающие на своей поверхности бактериальные токсины и часть патогенных микроорганизмов. Их применение – это симптоматическая терапия, позволяющая снизить уровень эндогенной интоксикации, уменьшить выраженность диареи и метеоризма.
- Противодиарейные средства.

Могут использоваться также витаминные комплексы, ферментные препараты, противоаллергические средства. Если же дисбактериоз развивается на фоне хронических заболеваний ЖКТ, подбирают лечение для их перевода в фазу стойкой ремиссии.
Куда обращаться
Лечить в СПб дисбактериоз кишечника могут врачи нескольких специальностей. Но все же профильным специалистом для этой патологии является гастроэнтеролог. Такой врач может рекомендовать необходимую схему обследования, грамотно оценить состояние всего желудочно-кишечного тракта, составить оптимальную программу лечения и дальнейшего восстановления. При выборе специалиста необходимо учитывать также профиль и возможности медицинского учреждения, по возможности отдавая предпочтение современным специализированным клиникам.
ICLINIC – это многопрофильный центр в г. Санкт-Петербург, ориентированный на достоверную диагностику и результативное лечение различных заболеваний органов пищеварения. В нашей клинике для обследования пациентов используется только современное высококлассное оборудование и прогрессивные диагностические программы, широко применяются эндоскопические методики.
В нашей клинике для обследования пациентов используется только современное высококлассное оборудование и прогрессивные диагностические программы, широко применяются эндоскопические методики.
Прием пациентов в ICLINIC ведут высококвалифицированные врачи различных специальностей, в штате имеется и сертифицированный опытный гастроэнтеролог. Лечение подбирается индивидуально, с учетом результатов обследования и современных клинических рекомендаций. Мы используем проверенные методики и результативные терапевтические схемы, добиваясь улучшения состояния даже у пациентов с тяжелыми и осложненными формами заболеваний.
Мы рекомендуем:
Прием врача-гастроэнтеролога
ФКС
ФКС с консультацией ведущего специалиста
Тест на вероятность рака желудка
Ваш возраст более 45 лет?
Да Нет
У Ваших родственников были онкологические заболевания?
Да Нет
У Вас есть хронические заболеваний желудочно-кишечного тракта:
— хронический гастрит,
— язвенная болезнь,
— хронический колит и другие воспалительные заболевания кишечника,
— болезнь Крона,
— неспецифический язвенный колит,
— выявленные ранее полипы желудка и кишечника,
— выявленные подслизистые эпителиальные образования желудочно-кишечного тракта?
Да Нет
Вы переносили операции на желудке и кишечнике?
Да Нет
У Вас есть рубцово-спаечные изменения желудочно-кишечного тракта?
Да Нет
Вы курите (более 1 сигареты в день)?
Да Нет
Вы допускаете погрешности в диете (низкое потребление овощей и фруктов, высокое потребление мяса и животных жиров)?
Да Нет
У Вас есть хотя-бы один из перечисленных симптомов:
— лишний вес,
— затруднения при глотании,
— раздражительность,
— бледность кожи,
— боли за грудиной,
— немотивированная слабость,
— нарушение сна,
— снижение аппетита,
— неприятный запах изо рта,
— отрыжка,
— тошнота и/или рвота,
— чувство тяжести в животе,
— изменение стула (запоры и/или поносы),
— следы крови в стуле,
— боли в животе.

Да Нет
Запишитесь на прием
В чем преимущества ICLINIC?Высочайший уровень специалистов: среди них доктора медицинских наук и члены мировых врачебных сообществ, а средний стаж врачей клиники – 16 лет безупречной работы.
Современное экспертное оборудование: диагностические аппараты медицинского центра выпущены в 2017 году ведущими мировыми производителями (Pentax и другими того же уровня).
Безупречная точность эндоскопической диагностики благодаря высокому разрешению изображения в 1,25 млн.пикселей.
Уникальные технологии ранней диагностики рака, среди которых i-scan – виртуальная хромоэндоскопия. С помощью данной технологии можно распознать даже самые мелкие, начальные опухолевые изменения.
Все для комфорта пациента: эффективное обезболивание, включая общий наркоз; тонкие эндоскопы менее 10мм в диаметре; быстрое и точное проведение манипуляций.
Безопасность: автоматизированная дезинфекция оборудования с контролем качества, мониторинг жизненно важных функций пациента в процессе исследований.
Узкая специализация: медцентр занимается заболеваниями пищеварительной системы, постоянно совершенствуясь именно в своей отрасли. Наши специалисты постоянно проходят повышение квалификации, участвуют в международных конференциях, тренингах и семинарах в России и Европе.
Удобное расположение: Петроградский район Санкт-Петербурга расположен недалеко от ценра. Сюда удобно добираться как на машине, так и на общественном транспорте. Совсем рядом с клиникой находится станция метро Чкаловская, также недалеко от медицинского центра станции СПб Спортивная, Петроградская и Горьковская.

Наш профессионализм всегда на страже вашего здоровья.
Возраст | Флора | Референсные значения |
Меньше 1 года | Бифидофлора | 1010 — 1011 Ед/л |
Молочнокислые бактерии | 106 — 108 Ед/л | |
Кишечная палочка с нормальными ферментативными свойствами | 107 — 108 Ед/л | |
Кишечная палочка с гемолизирующими свойствами | Отрицательно | |
Кишечная палочка со сниженными ферментативными свойствами | Не более 106 — 107 Ед/л | |
Кишечная палочка, лактозонегативные | 105 Ед/л и менее | |
Клостридии | Менее 103 Ед/л | |
Энтерококки | 105 — 107 Ед/л | |
Условно-патогенная флора (вся) | Менее 104 Ед/л | |
Staphylococcus aureus | Отрицательно | |
Дрожжевые грибы рода Candida | 103 и менее Ед/л | |
Candida albicans | 103 и менее Ед/л | |
Бактерии тифопаротифозной группы | Отрицательно | |
Бактерии дизентерийной группы | Отрицательно | |
ЭТ, ЭИ, ЭП эшерихии | Отрицательно | |
Больше 1 года | Бифидофлора | 109 — 1010 Ед/л |
Молочнокислые бактерии | 107 — 108 Ед/л | |
Кишечная палочка с нормальными ферментативными свойствами | 107 — 108 Ед/л | |
Кишечная палочка с гемолизирующими свойствами | Отрицательно | |
Кишечная палочка со сниженными ферментативными свойствами | Не более 107 — 108 Ед/л | |
Кишечная палочка, лактозонегативные | 105 Ед/л и менее | |
Клостридии | Менее 105 Ед/л | |
Энтерококки | 105 — 108 Ед/л | |
Условно-патогенная флора (вся) | Менее 104 Ед/л | |
Staphylococcus aureus | Отрицательно | |
Дрожжевые грибы рода Candida | 104 и менее Ед/л | |
Candida albicans | 104 и менее Ед/л | |
Бактерии тифопаротифозной группы | Отрицательно | |
Бактерии дизентерийной группы | Отрицательно | |
ЭТ, ЭИ, ЭП эшерихии | Отрицательно |
Сдать анализ на дисбактериоз с чувствительностью к бактериофагам в Москве
Анализ кала на чувствительность к бактериофагам необходим для количественного и качественного определения состава микрофлоры материала, предоставленного для анализа. В ходе данного обследования определяется патогенные и условно-патогенные микроорганизмы.
В ходе данного обследования определяется патогенные и условно-патогенные микроорганизмы.
На сегодняшний день микробиологическое исследование представляет собой наиболее информативный, раскрывающий полноту картины метод при инфекционных процессах. Способ дает возможность качественно и количественно охарактеризовать условно-патогенную микрофлору ЖКТ, выявить степень чувствительности к противомикробным препаратам отдельных представителей патогенных кишечных бактерий.
К условно-патогенным микроорганизмам ЖКТ относится подавляющее большинство представителей семейства Enterobacteriaceae: энтерококки негемолитического вида, стимулирующие выработку иммуноглобулинов и активно подавляющие разрастание численности патогенных бактерий, к которым относятся шигеллы и сальмонеллы, цитробактер, протей, энтеробактерии, клебсиеллы, стафилококки и др.
Выделяемые типы микроорганизмов и возбудители:
- патогенные виды энтеробактерий – шигеллы, сальмонеллы;
- УПМО (условно-патогенные микроорганизмы) – неферментирующие
грамотрицательные бактерии, стафилококки, энтеробактерии, энтерококки, грибы.

Что показывает анализ
При дисбактериозе кишечника происходит изменение его нормальной флоры на патогенную и условно-патогенную. В нормальном состоянии в кишечнике обитают грамположительные аэробные бактерии и грибы, среди которых преобладают следующие виды:
- молочнокислые палочки;
- стафилококки;
- стрептококки.
Такой симбиоз является совершенно нормальным явлением для организма. Соотношение между количеством микроорганизмов существенно не меняется с течением времени, за исключением случаев, когда уровень иммунитета падает и бифидобактерии, молочнокислые и кишечные палочки исчезают или их число значительно снижается. Им на смену приходят представители рода кандидов, стафилококки, протеи, синегнойные палочки.
Как проводится анализ
При определении имеющихся в кале бактерий проводят их посев на питательную среду. Затем определяется их видовая принадлежность.
При проведении исследования на чувствительность к фагам, для каждого типа колоний подбирают бумажные диски пропитанные препаратом, который оказывает воздействие на микроорганизмы.
UPM присоединяется к проекту под руководством BORDA, направленному на пилотирование решений в области санитарии для малообеспеченных сообществ в Иордании: UPM Umwelt-Projekt-Management GmbH |
UPM была выбрана в качестве одной из консалтинговых фирм для поддержки начального этапа и этапа планирования проекта InnovativeSanitation Solutions & Reuse in Arid Region (ISSRAR). Проект, инициированный Швейцарским агентством по развитию и сотрудничеству (SDC) и управляемый консорциумом BORDA — Seecon и под контролем Руководящего комитета, возглавляемого Управлением водного хозяйства Иордании, был запущен 6 декабря 2017 года. .
Проект направлен на повышение эффективности очистки сточных вод, улучшение управления фекальным осадком (FSM) и превращение потоков отходов в потоки физических и финансовых ресурсов путем обеспечения и содействия безопасному повторному использованию очищенных сточных вод и фекального осадка. Таким образом, проект будет способствовать снижению экологических рисков, созданию лучших условий жизни и повышению устойчивости принимающих сообществ в Иордании.
Таким образом, проект будет способствовать снижению экологических рисков, созданию лучших условий жизни и повышению устойчивости принимающих сообществ в Иордании.
Задачи группы экспертов UPM сосредоточены на предоставлении рекомендаций и изучении потенциальных объектов проекта, а также на высоком уровне мнений заинтересованных сторон и существующих технологических решений; вклад в выбор технологии и обзор предлагаемой системы лечения; концептуальные идеи для повторного использования ила / твердых биологических веществ; и разработка стратегии софинансирования санитарии в выбранной общине.
В области, где каждая капля воды имеет значение, новые модели сотрудничества и построения доверия между государственными структурами, предприятиями, сообществами и организациями гражданского общества позволяют внедрять устойчивые и жизнеспособные решения для решения проблемы нехватки воды. — Алекс Миллер, региональный директор BORDA в Западной и Центральной Азии и соруководитель проекта
— Алекс Миллер, региональный директор BORDA в Западной и Центральной Азии и соруководитель проекта
Мы очень гордимся возможностью поделиться знаниями и опытом UPM в области устойчивой и инновационной децентрализованной санитарии и решений FSM для этого захватывающего проекта в Иордании. Команда UPM уже работает в Иордании и очень рада быть частью местной и международной команды экспертов; вместе стремясь реализовать решения, основанные на сочетании требований сообщества и инновационных, но проверенных технических приложений. — Мартин Дилгер, управляющий директор UPM
Команда UPM уже работает в Иордании и очень рада быть частью местной и международной команды экспертов; вместе стремясь реализовать решения, основанные на сочетании требований сообщества и инновационных, но проверенных технических приложений. — Мартин Дилгер, управляющий директор UPM
Для получения дополнительной информации о проекте ISSRAR посетите веб-сайт BORDA West & Central Asia .
Для получения дополнительной информации о консультационных услугах UPM посетите наш веб-сайт https://www.upm-cdm.eu/ или позвоните по телефону:
Мартин Дилгер, управляющий директор, UPM Germany
mdilger (at) upm-cdm.eu, T: +49 89 1222197-50
или
Мари Рейссет, генеральный директор, UPM China
mreysset (at) upm-cdm.eu, T: +86 10 6468 0500
Сравнительная оценка микробного состава и метабонома фекалий свиней, фермеров и людей
Кау, А. Л., Ахерн, П. П., Гриффин, Н. В., Гудман, А. Л. и Джеффри, И. Питание человека, микробиом кишечника и иммунная система: видение будущего. Природа 474 , 327–336 (2012).
Артикул CAS Google Scholar
Исааксон Р. и Ким Х. Кишечный микробиом свиньи. Anim. Лечить. Res. Ред. 13 , 100–109 (2012).
Артикул Google Scholar
Marchesi, J. et al. . Микробиота кишечника и здоровье хозяина: новый клинический рубеж. Кишечник 65 , 330–339 (2016).
PubMed Статья Google Scholar
Каутари А. Э., Армугом Ф., Гордон Дж. И., Рауль Д. и Хенриссат Б. Изобилие и разнообразие углеводно-активных ферментов в микробиоте кишечника человека. Nat. Rev. Microbiol. 11 , 497–504 (2013).
PubMed Статья CAS Google Scholar

Вальдес, А. М., Уолтер, Дж., Сигал, Э. и Спектор, Т. Д. Роль кишечной микробиоты в питании и здоровье. BMJ 361 , 36–44 (2018).
Google Scholar
Rogers, G. B. et al. . От дисбактериоза кишечника до нарушения функции мозга и психических заболеваний: механизмы и пути. Мол. Психиатрия 21 , 738–748 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Кандела, М., Бьяджи, Э., Маккаферри, С., Туррони, С. и Бриджиди, П. Микробиота кишечника — это пластический фактор, реагирующий на изменения окружающей среды. Тенденции микробиологии 20 , 385–391 (2012).
CAS Статья Google Scholar
Миразо, С. и др. . Серологическое и вирусологическое исследование вируса гепатита Е (HEV) в резервуарах для животных в Уругвае выявило повышенную распространенность и очень тесную филогенетическую связь между линиями свиней и людей. Vet Microbiol 213 , 21–27 (2018).
Vet Microbiol 213 , 21–27 (2018).
CAS PubMed Статья Google Scholar
Song, S. et al. . Сожительствующие члены семьи разделяют микробиоту друг с другом и со своими собаками. Элиф 2 , e00458 (2013).
PubMed PubMed Central Статья Google Scholar
Pehrsson, E. et al. . Взаимосвязанные микробиомы и резистомы в местах обитания людей с низким доходом. Природа 533 , 212–216 (2016).
ADS CAS PubMed PubMed Central Статья Google Scholar
Вестергаард, Д. и др. . В домах свиноводов обитают более разнообразные сообщества переносимых по воздуху бактерий, чем в свинарниках или пригородных домах. Фронт. Microbiol. 9 , 1–14 (2018).
Артикул Google Scholar

Roduit, C., Frei, R., von Mutius, E. & R, L. Гипотеза гигиены. Влияние окружающей среды на иммунную систему (изд. Esser, C.) 77–96, https://doi.org/10.1007/978-3-7091-1890-0 (Springer, 2016).
Google Scholar
Тасним Н., Абулизи Н., Питер Дж., Харт М. и Гибсон Д. Связь микробной экосистемы кишечника с окружающей средой: зависит ли здоровье кишечника от того, где мы живем? Фронт. Microbiol. 8 , 1–8 (2017).
Артикул Google Scholar
Якобссон, Х. и др. . Кратковременное лечение антибиотиками оказывает различное долгосрочное воздействие на микробиом горла и кишечника человека. PLoS One 5 , e9836 (2010).
ADS PubMed PubMed Central Статья CAS Google Scholar
Ламенделла, Р., Доминго, Дж., Гош, С., Мартинсон, Дж. И Эртер, Д. Сравнительная метагеномика фекалий раскрывает уникальные функциональные возможности кишечника свиней. BMC Microbiol. 11 , 1–17 (2011).
Сравнительная метагеномика фекалий раскрывает уникальные функциональные возможности кишечника свиней. BMC Microbiol. 11 , 1–17 (2011).
Артикул CAS Google Scholar
Дойш, С., Тилокка, Б., Камаринья-Силва, А. и Зейферт, Дж. Новости исследований в области животноводства — Использование Omics-технологий для изучения микробиоты желудочно-кишечного тракта сельскохозяйственных животных. Comput. Struct. Biotechnol. J. 13 , 55–63 (2015).
CAS PubMed Статья Google Scholar
Ван, В., Ху, Х., Зийлстра, Р. Т., Чжэн, Дж. И Гензле, М. Г. Метагеномные реконструкции микробного метаболизма кишечника у поросят-отъемышей. Микробиом 7 , 1–11 (2019).
CAS Статья Google Scholar
Trinh, P., Zaneveld, J. R., Safranek, S. & Rabinowitz, P. M. Одно отношение к здоровью между людьми. микробиомов животных и окружающей среды: мини-обзор. Фронт. Публичное исцеление. 6 , 1–9 (2018).
M. Одно отношение к здоровью между людьми. микробиомов животных и окружающей среды: мини-обзор. Фронт. Публичное исцеление. 6 , 1–9 (2018).
Google Scholar
Smillie, C.С. и др. . Экология управляет глобальной сетью обмена генами, соединяющей человеческий микробиом. Природа 480 , 241–244 (2011).
ADS CAS PubMed PubMed Central Статья Google Scholar
Клоус, Г., Хасс, А., Хедерик, Д. Дж. Дж. И Коутиньо, Р. А. Контакты человека и домашнего скота и их связь с передачей зоонозных патогенов, систематический обзор литературы. Одно исцеление. 2 , 65–76 (2016).
Артикул Google Scholar
Tan, S., Chong, C., Teh, C., Ooi, P. & Thong, K. Возникновение вирулентных мультирезистентных Enterococcus faecalis и Enterococcus faecium у свиней, фермеров и фермерских хозяйств в Малайзия. PeerJ 6 , e5353 (2018).
PeerJ 6 , e5353 (2018).
PubMed PubMed Central Статья CAS Google Scholar
Ларсен, Дж. и др. . Устойчивый к гентамицину Enterococcus faecalis свиного происхождения у человека, Дания. Emerg. Заразить. Дис. 16 , 682–684 (2010).
PubMed PubMed Central Статья Google Scholar
Мессенджер, А. М., Барнс, А. Н. и Грей, Г. С. Обратная передача зоонозов (зооантропоноз): систематический обзор редко задокументированных биологических угроз для животных человеком. PLoS One 9 , 1–9 (2014).
Артикул CAS Google Scholar
Делахой, М. Дж. и др. . Патогены передаются с фекалиями животных в странах с низким и средним уровнем доходов. Внутр. J. Hyg. Environ. Здравоохранение 221 , 661–676 (2018).
PubMed PubMed Central Статья Google Scholar
Сан, Дж. и др. . Сравнение микробного состава фекалий и генов устойчивости к антибиотикам свиней, сельскохозяйственных рабочих и жителей окрестных деревень. Sci. Отчет 7 , 1–7 (2017).
ADS Статья CAS Google Scholar
Чонг, К. и др. . Влияние этнической принадлежности и социально-экономических различий на состав кишечной микробиоты у подростков в Малайзии. Sci. Отчет 5 , 1–12 (2015).
Google Scholar
Chae, J. P., Pajarillo, E. A. B., Oh, J. K., Kim, H. & Kang, D.-K. Выявление комбинированного воздействия лактулозы и пробиотических энтерококков на фекальную микробиоту свиней с помощью пиросеквенирования 454. Microb. Biotechnol. 9 , 486–495 (2016).
CAS PubMed PubMed Central Статья Google Scholar
Холман, Д., Брунель, Б., Трахсел, Дж. И Аллен, Х. Мета-анализ для определения основной микробиоты в кишечнике свиней. mSystems 2 , e00004–17 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Гринхал, К., Мейер, К. М., Аагард, К. М. и Уилмс, П. Микробиом кишечника человека в здоровье: формирование и устойчивость микробиоты на протяжении всей жизни. Environ.Microbiol. 18 , 2103–2116 (2016).
PubMed Статья Google Scholar
Walsh, H., Haq, H., Cersosimo, L., Kien, C. L. и Kraft, J. Снижение численности Firmicutes в микробиоте кишечника после употребления диеты, содержащей молочные жиры. FASEB J. 30 , 683.11 (2016).
Google Scholar
Лю Б. и др. . Ответ микробиоты кишечника на пищевые волокна и метаболическое взаимодействие с SCFAs у поросят.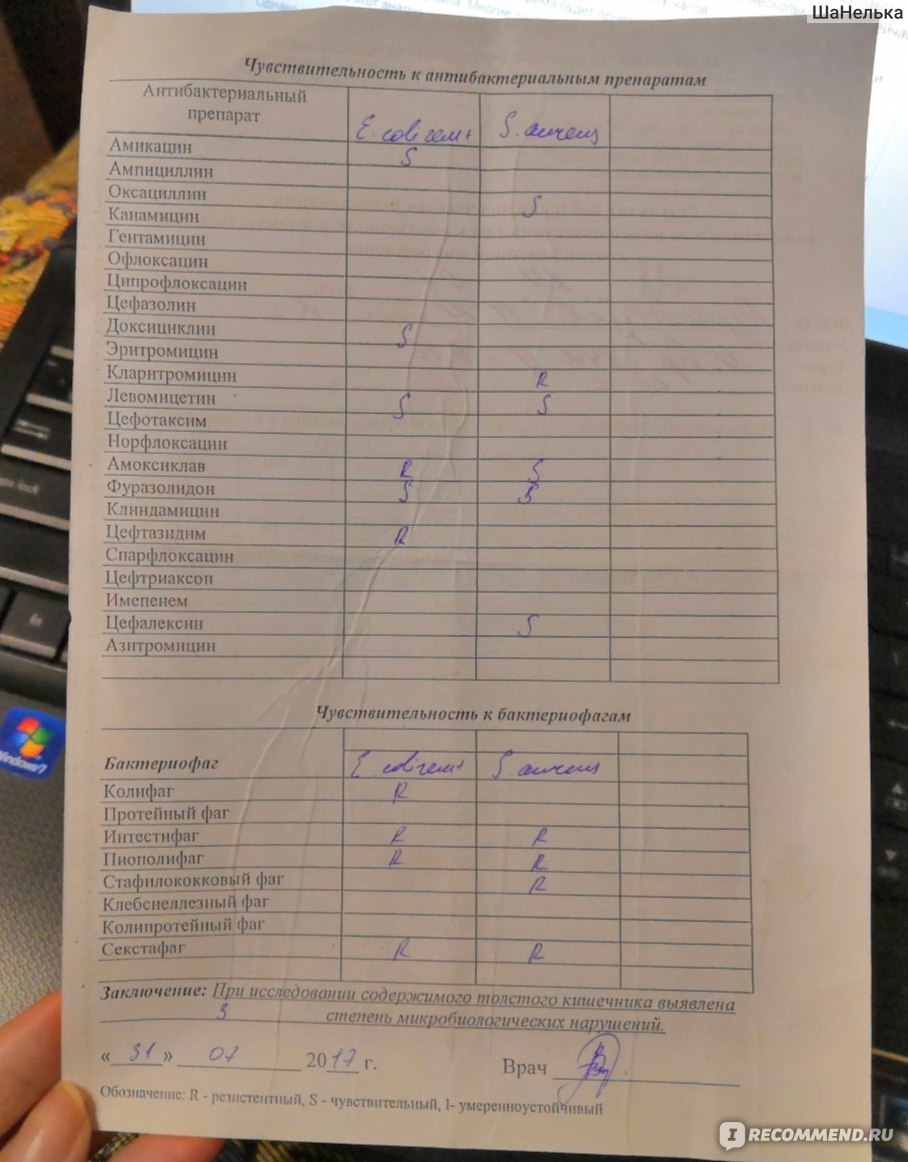 Фронт. Microbiol. 9 , 1–12 (2018).
Фронт. Microbiol. 9 , 1–12 (2018).
Артикул Google Scholar
Мариат Д. и др. . Соотношение фирм и бактероидов в микробиоте человека меняется с возрастом. BMC Microbiol. 9 , 1–6 (2009).
Артикул CAS Google Scholar
Лей, Р., Тернбо, П., Кляйн, С. и Гордон, Дж. Микробы кишечника человека, связанные с ожирением. Природа 444 , 1022–1023 (2006).
ADS CAS PubMed Статья Google Scholar
Коляда А. и др. . Связь между индексом массы тела и соотношением Firmicutes / Bacteroidetes у взрослого населения Украины. BMC Microbiol. 17 , 1–6 (2017).
Артикул CAS Google Scholar
Го, Х. и др. . Разработка метода ПЦР в реальном времени для Firmicutes и Bacteroidetes в фекалиях и его применение для количественной оценки кишечной популяции тучных и постных свиней. Lett. Прил. Microbiol. 47 , 367–373 (2008).
Lett. Прил. Microbiol. 47 , 367–373 (2008).
CAS PubMed Статья Google Scholar
Морено, J. Prevotella copri и микробный патогенез ревматоидного артрита. Reumatol. Clin. 11 , 61–63 (2015).
PubMed Статья Google Scholar
Маеда Ю. и Такеда К. Роль кишечной микробиоты при ревматоидном артрите. J. Clin. Med. 6 , 60 (2017).
PubMed Central Статья CAS Google Scholar
Miquel, S. et al. . Faecalibacterium prausnitzii и здоровье кишечника человека. Curr. Opin. Microbiol. 16 , 255–261 (2013).
CAS PubMed Статья Google Scholar
Феррейра-Хальдер, К. В., Фариа, А. В., де, С. и Андраде, С. С. Действие и функция Faecalibacterium prausnitzii в отношении здоровья и болезней. Best Pract. Res. Clin. Гастроэнтерол. 31 , 643–648 (2017).
Best Pract. Res. Clin. Гастроэнтерол. 31 , 643–648 (2017).
CAS PubMed Статья Google Scholar
Эппинга, Х. и др. . Подобное истощение защитных Faecalibacterium prausnitzii при псориазе и воспалительном заболевании кишечника, но не при гнойном гидрадените. Колит Дж. Крона 10 , 1067–1075 (2016).
Артикул Google Scholar
Лопес-Силес, М., Дункан, С. Х., Гарсия-Хиль, Л. Дж. И Мартинес-Медина, М. Faecalibacterium prausnitzii: От микробиологии к диагностике и прогнозированию. ISME J. 11 , 841–852 (2017).
PubMed PubMed Central Статья Google Scholar
Quévrain, E. et al. . Идентификация противовоспалительного белка Faecalibacterium prausnitzii, комменсальной бактерии, дефицитной при болезни Крона. Кишечник 65 , 415–425 (2016).
PubMed Статья CAS Google Scholar
Rinkinen, M. L., Koort, J. M. K., Ouwehand, A. C., Westermarck, E. & Björkroth, K. J. Streptococcus alactolyticus — доминирующие культивируемые виды молочнокислых бактерий в тонкой кишке и фекалиях четырех собак со свищами. FEMS Microbiol. Lett. 230 , 35–39 (2004).
CAS PubMed Статья Google Scholar
Девризе, Л. А., Хоммез, Дж., Пот, Б. и Хезебрук, Ф. Идентификация и состав стрептококковой и энтерококковой флоры миндалин, кишечника и фекалий свиней. J. Appl. Бактериол. 77 , 31–36 (1994).
CAS PubMed Статья Google Scholar
Almeida, P., Railsback, J. & Gleason, J. B. Редкий случай инфекционного эндокардита Streptococcus alactolyticus, осложненного септической эмболой и микотической аневризмой левой средней мозговой артерии. Case Rep. Infect. Дис. 2016 , 1–3 (2016).
Case Rep. Infect. Дис. 2016 , 1–3 (2016).
Google Scholar
Топфнер, Н. и др. . Фульминантный неонатальный сепсис, вызванный Streptococcus alactolyticus — отчет и обзор случая. Apmis 122 , 654–656 (2014).
PubMed Статья Google Scholar
Шенг, К. К., Янг, З. Дж., Чжао, Х. Б., Ван, X. Л. и Го, Дж. Ф. Влияние L-триптофана, фруктана и казеина на снижение содержания аммиака, сероводорода и скатола в ферментированном свином навозе. Азиатско-австралийский. J. Anim. Sci. 28 , 1202–1208 (2015).
CAS Google Scholar
Николсон, Дж. К. и др. . Метаболические взаимодействия микробиоты кишечника и хозяина. Наука (80-.). 108 , 1262–1268 (2012).
ADS Статья CAS Google Scholar

Ден Аббеле, П. В. и др. . Производящие бутират виды Clostridium cluster XIVa специфически колонизируют муцины в модели кишечника in vitro . ISME J. 7 , 949–961 (2013).
PubMed Статья CAS Google Scholar
Мейер, К., Де Вос, П. и Прибе, М. Г. Бутират и другие короткоцепочечные жирные кислоты как модуляторы иммунитета: какое значение для здоровья? Curr. Opin. Clin. Nutr. Метаб. Уход 13 , 715–721 (2010).
CAS PubMed Статья Google Scholar
Коэт, Р. А. и др. . Метаболизм L-карнитина, питательного вещества красного мяса, микробиоты кишечника способствует развитию атеросклероза. Nat. Med. 19 , 576–585 (2013).
ADS CAS PubMed PubMed Central Статья Google Scholar
Zhu, W., Zeneng, W.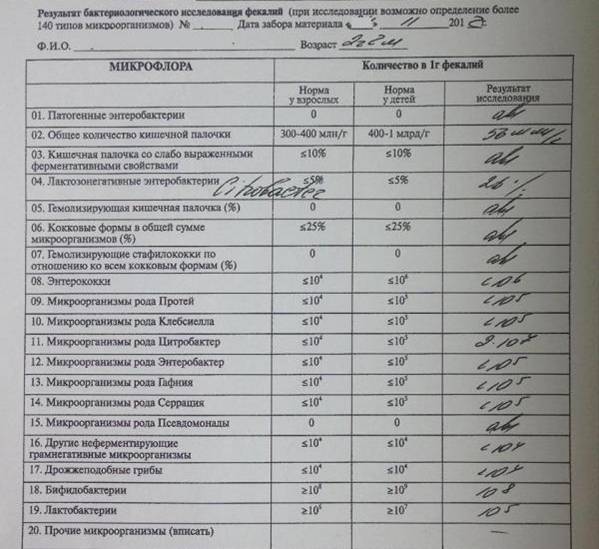 , Tang, W. H. W. & Hazen, S. L. ТМАО, генерируемое кишечными микробами из пищевого холина, является протромботическим у субъектов. Тираж 135 , 1671–1673 (2017).
, Tang, W. H. W. & Hazen, S. L. ТМАО, генерируемое кишечными микробами из пищевого холина, является протромботическим у субъектов. Тираж 135 , 1671–1673 (2017).
PubMed PubMed Central Статья Google Scholar
Pianta, A. et al. . Доказательства иммунной значимости кишечного микроба Prevotella copri у пациентов с ревматоидным артритом. Arthritis Rheumatol. 69 , 964–975 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Баджер, Л. и др. . Отчетливые профили микробиоты кишечника у пациентов с первичным склерозирующим холангитом и язвенным колитом. World J. Gastroenterol. 23 , 4548–4558 (2017).
CAS PubMed PubMed Central Статья Google Scholar
Бруска, С. Б., Абрамсон, С. Б. и Шер, Дж. У. Микробиом и воспаление слизистой оболочки как внесуставные триггеры ревматоидного артрита и аутоиммунитета. Curr. Opin. Ревматол. 26 , 101–107 (2014).
У. Микробиом и воспаление слизистой оболочки как внесуставные триггеры ревматоидного артрита и аутоиммунитета. Curr. Opin. Ревматол. 26 , 101–107 (2014).
CAS PubMed PubMed Central Статья Google Scholar
Scher, J. U. et al. . Расширение кишечной Prevotella copri коррелирует с повышенной восприимчивостью к артриту. Elife 2 , 1–20 (2013).
Артикул CAS Google Scholar
Чонг, К. В. и др. . Влияние окружающей среды на бактериальное разнообразие почв на острове Сигни в морской Антарктике. Polar Biol. 32 , 1571–1582 (2009).
Артикул Google Scholar
Schloss, P. D. et al. . Представляем mothur: программное обеспечение с открытым исходным кодом, независимое от платформы, поддерживаемое сообществом, для описания и сравнения сообществ микробов. Заявл. Environ. Microbiol. 75 , 7537–7541 (2009).
CAS PubMed PubMed Central Статья Google Scholar
Рохарт, Ф., Готье, Б., Синг, А. и Ле Цао, К. A. mixOmics: пакет R для выбора функций omics и интеграции нескольких данных. PLoS Comput. Биол. 13 , e1005752 (2017).
ADS PubMed PubMed Central Статья CAS Google Scholar
Лав, М. И., Хубер, В. и Андерс, С. Умеренная оценка кратного изменения и дисперсии данных РНК-секвенирования с помощью DESeq. 2. Genome Biol. 15 , 550 (2014).
PubMed PubMed Central Статья CAS Google Scholar
Полная последовательность генома EC1-UPM, нового N4-подобного бактериофага, который инфицирует Escherichia coli O78: K80
Резюме
Предпосылки
Бактериофаг EC1-UPM представляет собой N4-подобный бактериофаг, который специфически заражает Escherichia coli O78: K80, патогенный штамм птиц, вызывающий колибактериоз у домашней птицы.Полная последовательность генома бактериофага EC1-UPM была проанализирована и сравнена с другими тесно связанными группами N4-подобных фагов для оценки их генетического сходства и различий.
Результаты
Бактериофаг EC1-UPM имеет очень похожий профиль использования кодонов со своим хозяином и не содержит гена тРНК. Сравнительный геномный анализ показывает близкое сходство бактериофага EC1-UPM с тремя N4-подобными бактериофагами, а именно vB_EcoP_G7C, IME11 и KBNP21, с 44 генами, кодирующими белок, с общим порогом идентичности 70%.Геномная область, кодирующая белок хвостового волокна, оказалась уникальной для бактериофага EC1-UPM. Дальнейшее аннотирование белка хвостового волокна с помощью HHpred, высокочувствительного инструмента определения гомологии, выявляет присутствие структуры белка, гомологичной различным белкам, обрабатывающим полисахариды, на его С-конце. Используя доступность нескольких последовательностей генома N4-подобных бактериофагов, были идентифицированы основные гены N4-подобных бактериофагов и использованы для выполнения многолокационного филогенетического анализа, который позволил построить филогенетическое дерево с большей достоверностью, чем филогенетические деревья, основанные на отдельных генах.
Заключение
Мы впервые сообщаем о полной последовательности генома N4-подобного бактериофага, литического против патогенного птичьего патогена Escherichia coli O78: K80. Новый белок хвостового волокна с 928 аминокислотными остатками был идентифицирован в EC1-UPM, который может быть полезен для дальнейшего понимания специфичности фага-хозяина. Мультилокусный филогенетический анализ с использованием коровых генов секвенированных N4-подобных фагов показал, что эволюционные отношения хорошо коррелируют с паттерном специфичности хозяина.
Ключевые слова: Бактериофаг EC1-UPM, белок хвостовых волокон, полный геном, мультилокусный филогенетический анализ вызывает колибактериоз кур, индеек и других видов птиц любого возраста. Инфекция, связанная с респираторной инфекцией (аэросаккулит), за которой следует перигепатит, перикардит и сепсис, является разрушительной инфекцией, поскольку она может снизить рост и яйценоскость домашней птицы.В необработанных условиях регистрируется высокий уровень смертности птиц, что влечет за собой большие экономические потери для фермеров и птицеводства [1]. Эти бактерии попадают в пищевую цепочку человека через загрязнение окружающей среды фекальными пометами инфицированных цыплят и загрязненными продуктами из птицы [2]. В последние годы сообщалось, что обычное лечение с использованием антибиотиков менее эффективно из-за появления у возбудителя устойчивости к антибиотикам [3].
В наших предыдущих исследованиях бактериофаг EC1-UPM, выделенный из образца фекалий цыплят, способен снижать тяжесть инфекции, вызванной E.coli O78: K80 и потенциально может использоваться для лечения колибактериоза у кур. Основываясь на результатах нашего исследования in vivo , общая смертность цыплят снизилась на 70%, когда инфицированных цыплят лечили бактериофагом EC1-UPM. Масса тела обработанных цыплят была на 15,4% выше, чем у необработанных цыплят [4,5]. Для дальнейшего использования потенциала бактериофага EC1-UPM важно иметь представление о его генетической структуре, особенно о генах, которые ответственны за инфицирование и лизис бактерий-хозяев.
В этом исследовании мы впервые сообщаем о геноме бактериофага EC1-UPM, N4-подобного бактериофага, который инфицирует E. coli O78: K80. Генетические компоненты бактериофага EC1-UPM, которые могут отвечать за его специфичность и вирулентность, были идентифицированы и проанализированы in-silico . Мы также идентифицировали основные гены из доступных геномов N4-подобных бактериофагов и использовали эту информацию, чтобы получить представление об эволюционных взаимоотношениях N4-подобных бактериофагов с использованием мультилокусного филогенетического анализа.
Результаты и обсуждение
Геномные особенности бактериофага EC1-UPM
Геном бактериофага EC1-UPM состоит из 70 912 п.н. (содержание GC 42,9%). Всего было предсказано 80 генов, кодирующих белок, со средней длиной 816 п.н. (плотность кодирования 92,5%). Аннотация большинства предсказанных CDS показала уверенные попадания в белки N4-подобных бактериофагов, что указывает на то, что бактериофаг EC1-UPM может быть членом N4-подобных бактериофагов (дополнительный файл 1: Таблица S1).N4-подобные бактериофаги являются бактериофагами семейства Podoviridae . Члены этой группы настроены против своих хозяев. Специфичность N4-подобных бактериофагов к хозяину весьма разнообразна: от энтеробактерий, таких как E. coli и Pseudomonas aeruginosa , до морских бактерий, таких как Sulfitobacter sp. EE-36 и Silicibacter pomeroyi DSS-3 [6-9]. Бактериофаг N4, поражающий E. coli K-12, в настоящее время является наиболее изученным штаммом этой группы бактериофагов [10].Помимо полного секвенирования его генома, идентичность и расположение нескольких его структурных белков были установлены путем сравнения трехмерных криоэлектронно-микроскопических структур N4 дикого типа и его мутантов [11]. На основании нашего анализа аннотированные белки бактериофага EC1-UPM можно разделить на следующие функциональные группы: структура и упаковка бактериофага (портальный белок, основной белок оболочки, хвостовой белок, белок хвостовой шишки, структурный белок, белок, украшающий капсид), репликация ДНК. / модификация (ДНК-геликаза, ДНК-полимераза, эндонуклеаза, терминаза), трансдукция сигнала и регуляторная функция (ssDNA-связывающий белок, РНК-полимераза), метаболизм нуклеотидов (тимидилатсинтаза, dCTP дезаминаза) и лизис хозяина (холин, N-ацетилмурамидаза).Присутствие гена лизиса, но не гена, связанного с лизогенией, указывает на то, что бактериофаг EC1-UPM является литическим бактериофагом. Самый большой ген кодирует полимеразу вириона и составляет 10 839 п.н., что составляет примерно 15% от всей длины генома (рисунок). Наличие кодируемой бактериофагом РНК-полимеразы такого размера является признаком бактериофага N4. На бактериофаге N4 было продемонстрировано, что эта особенно большая РНК-полимераза была упакована в его капсид и выброшена в клетку-хозяин, что устраняет необходимость полагаться на РНК-полимеразу хозяина для транскрипции его ранних генов [11].Никакая тРНК не была идентифицирована в геноме бактериофага EC1-UPM, что указывает на то, что при попадании в организм хозяина он полностью зависит от состава тРНК хозяина для синтеза белка.
Линейная визуализация генома бактериофага EC1-UPM. Направление стрелки соответствует ориентации транскрипции. HP, гипотетический белок; PTF, фактор транскрипции, связанный с патогенезом; DNS, внеклеточная дезоксирибонуклеаза.
Использование кодонов бактериофага EC1-UPM
Сравнение использования кодонов в E.coli O78: K80 и бактериофаг EC1-UPM выявили довольно похожую частоту кодонов (рисунок). Совместимость использования кодонов фага с кодоном его хозяина даст эволюционное преимущество, поскольку это поможет в энергоемком и трудоемком механизме синтеза белка. Однако три кодона были чрезмерно представлены в бактериофаге EC1-UPM, а именно кодоны ACT, GTA и GCT, которые соответствуют аминокислотам треонину, валину и аланину, соответственно. Интересно, что бактериофаг EC1-UPM не адаптируется к различиям в использовании кодонов, о чем свидетельствует отсутствие гена тРНК в его геноме.В процессе эволюции фагов дцДНК присутствие тРНК, особенно тех, которые в изобилии в фаге и менее распространены в хозяине, позволяет фагу опередить своих потенциальных конкурентов в хозяине, более эффективно транслируя свои белки, ускоряя скорость размножения и снижая его время ожидания [12]. В фагах Т4, делеция генов тРНК, как было показано, снижает размер взрыва и скорость синтеза белка, демонстрируя ростовое преимущество при переносе генов тРНК в фагах [13]. Полное отсутствие тРНК в бактериофаге EC1-UPM может означать, что его способность инфицировать E.coli O78: K80 мог быть недавним приобретением.
Частота кодонов бактериофага EC1-UPM и его хозяина E. coli O78: K80. Буквы без скобок на оси X представляют аминокислоту, связанную с кодоном.
Сравнительная геномика бактериофага EC1-UPM и близкородственных N4-подобных бактериофагов
Первоначальный полный геномный BLAST бактериофага EC1-UPM по базе данных NCBI показал, что бактериофаг EC1-UPM тесно связан с тремя N4-подобными вирусами, а именно с KBNP21, vB_EcoP_G7C и IME11 (данные не показаны).Бактериофаги KBNP21, vB_EcoP_G7C и IME11 были выделены из куриной фермы, конских фекалий и больничных сточных вод соответственно [6,14,15]. Бактериофаг IME11 может инфицировать 13 из 31 энтеропатогенного клинического штамма E. coli [6]. Бактериофаг vB_EcoP_G7C может инфицировать штамм 4 E. coli , тогда как бактериофаг KBNP21 может инфицировать штаммов E. coli KBP21 и KBP135 [14,15]. Однако отсутствие данных об антигенах O и K для тестируемых штаммов E. coli серьезно затрудняло определение специфичности хозяина на основе поверхностного антигена.Используя генератор изображений Blast Ring (BRIG), было показано, что большинство геномов консервативны в четырех бактериофагах (рисунок), за исключением области генома, включающей гены, кодирующие белки хвостовых волокон и шипов хвоста. Белки хвостовых волокон и шипов хвоста часто связаны со связыванием и деградацией поверхностного антигена клетки-хозяина [14]. Учитывая различия в источниках выделения каждого E. coli -инфицирующего N4-подобного бактериофага, вероятно, что их E.coli принадлежат к разным штаммам, у которых отсутствует сходство по профилю поверхностного антигена. Ген, кодирующий белок шипа на хвосте бактериофага EC1-UPM, консервативен только в бактериофаге KBNP21, который аналогичным образом был выделен из птичьего источника. Хотя аннотация InterProscan белка шипа хвоста не выявила какого-либо значимого белкового домена (дополнительный файл 1: таблица S1), увеличение аннотации белка с помощью HHpred указывает на три основных домена, связанных со связыванием и деградацией углеводов между остатками 120-190, 230 до 500 и от 550 до 700 (рисунок).Было показано, что остроконечный хвост является жизненно важным компонентом, особенно при инфекции бактерий, обладающих как O-, так и K-антигенами [14]. Некоторые K-антигены могут препятствовать прямому распознаванию рецептора на O-антигене фаговым хвостом из-за большого размера K-антигена (до 4000 A). Белок шипа хвоста функционирует, чтобы преодолеть такое ограничение, создавая туннель в капсуле клетки и открывая рецептор для белка хвоста [15,16].
Изображение бластного кольца с использованием полного генома бактериофага EC1-UPM в качестве эталона. Наружные кольца представляют собой геномы трех N4-подобных бактериофагов, тесно связанных с EC1-UPM.
Идентификация участков белка, гомологичных ферментам, расщепляющим полисахариды, в белках хвостовых шипов и хвостовых волокон бактериофага EC1-UPM с помощью HHpred. Идентификаторы белков указаны в квадратных скобках.
Новая область на С-конце белка хвостового волокна
На основании визуализации BRIG геномная область, соответствующая С-концу белка хвостового волокна бактериофага EC1-UPM, не была общей ни для одного члена близкородственного N4-подобного бактериофаги (рисунок).Подобно белку шипа хвоста, волокно хвоста бактериофага также участвует в специфичности хозяина. Сообщалось, что С-конец белка хвостового волокна изменяет круг хозяев бактериофага [17]. Таким образом, расхождения на С-конце могут отражать адаптацию фага к огромному разнообразию антигенов О и К в E. coli [18]. Расширение поиска BLAST на всю базу данных NR не выявило значительного попадания в область C-конца. На основании аннотации HHpred, C-конец белка хвостового волокна продемонстрировал значительную степень структурной гомологии с различными ферментами, разрушающими полисахариды, обладающими правой структурой β-спирали (рисунок и дополнительный файл 2: таблица S2).Примечательно, что остатки с 402 по 731 хвостового волокна демонстрируют значительное сходство с экзо-поли-α-D-галактуронозидазой, содержащей структуру β-спирали, у термофильных бактерий Thermotoga maritima, a (значение E = 4,44 × 10 -44 ) (рисунок и дополнительный файл 2: таблица S2). Экзо-поли-α-D-галактуронозидаза участвует в гидролизе пектиновой кислоты из невосстанавливающих с высвобождением дигалактуроната. Первоначально идентифицированная в пектатлиазе C, правая β-спираль обычно обнаруживается в каталитических доменах различных белков, которые обрабатывают олиго- и полисахариды [19].Учитывая структурную гомологию С-конца хвостового волокна с различными ферментами, разрушающими полисахариды (дополнительный файл 2: таблица S2), вполне вероятно, что хвостовое волокно может быть связано со связыванием и / или деградацией полисахарида, присутствующего на бактериальных антигенах. Присутствие гидролитических белков хвоста фага, как было показано, позволяет бактериофагу ΦK1-5 присоединяться и разрушать капсульные полисахариды K1 и K5 [20]. Дальнейшие ферментативные исследования необходимы для подтверждения функции белка хвостовых волокон бактериофага EC1-UPM и диапазона его субстратов.Этот новый белок хвостового волокна состоял из 928 аминокислотных остатков с молекулярной массой и pI 99,3 кДа и 4,85 соответственно.
Лизис хозяина
Широко признано, что лизис хозяина необходим для высвобождения и диспергирования потомков всех бактериофагов, за исключением нитчатых бактериофагов [21,22]. В бактериофаге EC1-UPM гены, участвующие в системе холин-эндолизин, расположены рядом друг с другом. N-ацетилмурамидаза (регистрационный номер NCBI: {«type»: «entrez-protein», «attrs»: {«text»: «AGC31576», «term_id»: «441431492», «term_text»: «AGC31576»}} AGC31576), который может быть ответственным за активность эндолиза, содержит домен, связанный с гликозилгидролазой (PF05838), от остатков с 22 до 113.Семейство гликозилгидролаз функционирует посредством гидролиза связи β-1,4 между N-ацетилмурамовой кислотой и N-ацетилглюкозамином в пептидогликановом слое бактерий. Присутствие пептидогликанового связывающего домена (PF09374) от остатка 116 до 195 после домена гликозилгидролазы может дополнительно усиливать активность N-ацетилмурамидазы за счет локализации белка в его целевой области. Визуализация топологии мембраны холинового белка бактериофага EC1-UPM показывает, что он содержит два трансмембранных домена с N- и C-концом (данные не показаны), расположенными в цитоплазматической области, таким образом классифицируя его как холин класса II [21,22].Гены, кодирующие систему холин-эндолизин, хорошо законсервированы в бактериофагах KBNP21, vB_EcoP_G7C, IME11 и EC1-UPM (рисунок), что указывает на общий механизм лизиса хозяина.
Идентификация ортологов и мультилокусный филогенетический анализ N4-подобных бактериофагов
Недавнее устойчивое увеличение и доступность полных геномов различных N4-подобных бактериофагов в GenBank дает возможность идентифицировать ортологи, которые хорошо сохраняются во всех N4- как бактериофаги.Первоначальная идентификация ортологов в бактериофаге EC1-UPM и его близкородственных штаммах vB_EcoP_G7C, IME11 и KBNP21 показала, что при 70% идентичности и 70% выровненных пороговых значениях в бактериофагах сохранялось в общей сложности 44 белка (таблица). Средняя идентичность ортологов, общих у бактериофагов vB_EcoP_G7C, IME11 и KBNP21, по сравнению с EC1-UPM составила 91%, 93% и 95% соответственно. Никакие ортологи не могут быть идентифицированы, когда анализ был расширен за счет включения более отдаленных N4-подобных бактериофагов (таблица) с использованием аналогичных настроек.Тем не менее, при снижении порога идентичности до 40% были идентифицированы семь ортологичных кластеров, а именно ДНК-полимераза I, N4 gp42-подобный белок, ДНК-примаза, N4 gp44-подобный белок, портальный белок, терминаза и N4 gp69-подобный белок. Функциональное назначение белков в ортологичных кластерах согласуется с представлением о том, что вирусы имеют общие гомологичные «гены-признаки», кодирующие белки, участвующие в репликации генома и формировании структуры вириона [23]. Анализ филогенетического дерева с использованием обрезанного и конкатенированного выравнивания белков в этих ортологичных кластерах показал, что эволюционное родство N4-подобных бактериофагов очень сильно коррелирует со специфичностью их хозяина.Сравнение нескольких филогенетических деревьев, построенных с использованием отдельных ортологичных кластеров с известной функцией (ДНК-полимераза, терминаза, портальный белок и ДНК-примаза), показало, что анализ филогенетического дерева, основанный исключительно на выравнивании последовательностей ДНК-полимеразы, привел к немного другой топологии дерева с более низкой поддержкой начальной загрузки. по сравнению с многолокусным филогенетическим деревом (рисунок). Это демонстрирует, что, хотя ДНК-полимераза является хорошим индикатором эволюционной взаимосвязи в целом, следует соблюдать осторожность при разрешении близкородственных N4-подобных бактериофагов.Напротив, построение филогенетического дерева на основе ДНК-примазы очень сравнимо с подходом многолокусной филогении с точки зрения топологии дерева и ценности поддержки бутстрапа.
Таблица 1
Ортологическая группа белков, консервативных для бактериофагов EC1-UPM, vB_EcoP_G7C (E), IME11 (I) и KBNP21 (K), как определено PanOCT (идентичность> 70% и выровненная длина> 70%)
| Идентификатор белка | Описание продукта | % Идентификатор | InterProscan ID | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| E | гипотетический белок | 87 | 95 | 97 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31513», «term_id»: «441431429», «term_text» : «AGC31513»}} AGC31513 | РНК-полимераза RNAP1 субъединица A | 9 6 | 94 | 98 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31517», «term_id»: «441431433», » term_text «:» AGC31517 «}} AGC31517 | гипотетический белок | 94 | 93 | 96 | ||||||
| {» type «:» entrez -rs- «text»: «AGC31519», «term_id»: «441431435», «term_text»: «AGC31519»}} AGC31519 | предполагаемый фаговый белок | 74 | 90 | 84 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31522», «term_id»: «441431438», «term_text»: «AGC31522»}} AGC31522 | гипотетический белок | 82 | 99 | 99 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31523», «term_id»: «441431439» , » term_text «:» AGC31523 «}} AGC31523 | гипотетический белок | 91 | 88 | 91 | ||||||
| {» type «:» entrez -rs- «text»: «AGC31525», «term_id»: «441431441», «term_text»: «AGC31525»}} AGC31525 | Субъединица B РНК-полимеразы | 95 | 96 | 99859 | 99859 908 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31526», «term_id»: «441431442», «term_text»: «AGC31526»}} AGC31526 | РНК-полимераза субъединица A | 88 | 96 | 97 | IPR002092 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31527», «term» : «441431443», «term_text»: «AGC31527»}} AGC31527 | Белок для украшения капсида | 79 | 91 | 93 | 9085 7 IPR003599; IPR007110; IPR013783||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31528», «term_id»: «441431444», «term_text»: «AGC31528»}} AGC31528 | гипотетический белок | 98 | 97 | 97 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31531», «term_id» : «441431447», «term_text»: «AGC31531»}} AGC31531 | гипотетический белок | 95 | 97 | 95 | ||||||
| » {тип entre « | , «attrs»: {«text»: «AGC31533», «term_id»: «441431449», «term_text»: «AGC31533»}} AGC31533 | гипотетический белок | 95 | 95 | IPR027417 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31534», «term_id»: «441431450», «term_text»: «AGC31534»}} AGC31534 | hy потетический белок | 95 | 97 | 99 | IPR018698; IPR025154 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31537», «term_id»: «441431453», «term_text»: «AGC31537»}} AGC31537 | протеин предполагаемого мембранного иммунитета | 86 | 98 | 94 | IPR016410 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC3 «term_id»: «441431455», «term_text»: «AGC31539»}} AGC31539 | гипотетический белок | 88 | 94 | 95 | 1 | тимидилатсинтаза | 84 | 95 | IPR003669 | |
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31543», «term_id»: «441431459», «term_text»: «AGC31543» }} AGC31543 | гипотетический белок | 76 | 80 | 82 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31545» term_id «:» 441431461 «,» term_text «:» AGC31545 «}} AGC31545 | rIIA-подобный белок | 86 | 91 | 94 | IPR003594 тип | IPR003594 «entrez-protein», «attrs»: {«text»: «AGC31546», «term_id»: «441431462», «term_text»: «AGC31546»}} AGC31546 | rIIB-подобный белок | 88 | 89 | 87 |
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31547», «term_id»: «441431463», «term_text»: «AGC31547»}} AGC31547 | гипотетический белок | 90 | 96 | 95 | ||||||
| {«type»: «entr ez-protein «,» attrs «: {» text «:» AGC31549 «,» term_id «:» 441431465 «,» term_text «:» AGC31549 «}} AGC31549 | ДНК-геликаза | 98 | 99 | 99 | IPR027417; IPR027785 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31550», «term_id»: «441431466», «term_text»: «AGC31550»}} AGC31550 | гипотетический белок | 97 | 99 | 98 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31551», «term_id» : «441431467», «term_text»: «AGC31551»}} AGC31551 | ДНК-полимераза | 98 | 98 | 97 | IPR001098; IPR002298; IPR012337 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31552», «term_id»: «441431468», «term_text»: «AGC31552»}} AGC31552 | гипотетический белок | 92 | 91 | 99 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31555», «term_id» : «441431471», «term_text»: «AGC31555»}} AGC31555 | гипотетический белок | 97 | 95 | 98 | ||||||
| » {тип entrez | » , «attrs»: {«text»: «AGC31557», «term_id»: «441431473», «term_text»: «AGC31557»}} AGC31557 | Белок DNS | 97 | 33 96 | 100 908 | IPR014820 | ||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31558», «term_id»: «441431474», «term_text»: «AGC31558»}} AGC31558 | гипотетический al протеин | 99 | 99 | 99 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31559», «term_id»: «441431475», «term_text»: «AGC31559»}} AGC31559 | оцДНК-связывающий белок | 95 | 97 | 97 | ||||||
| гипотетический белок | 99 | 99 | ||||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31561», «term_id»: «441431477», «term_text»: «AGC31561»}} AGC31561 | гипотетический белок | 76 | 92 | 94 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AG C31567 «,» term_id «:» 441431483 «,» term_text «:» AGC31567 «}} AGC31567 | предполагаемый структурный белок 88 | 85 | 85 | гипотетический белок | 92 | 93 | 94 | |||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31569», «term_id»: «441431485», «term_text»: «AGC31569»}} AGC31569 | предполагаемый хвостовой белок | 94 | 82 | 96 | ||||||
| {«type»: «entrez-protein»: «text» «:» AGC31570 «,» term_id «:» 441431486 «,» term_text «:» AGC31570 «}} AGC31570 | гипотетический белок | 96 | 97 | 99 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31571», «term_id»: «441431487», «term_text»: «AGC31571»}} AGC31571 | главный белок оболочки | 97 | 97 | 100 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31572» «term_id»: «441431488», «term_text»: «AGC31572»}} AGC31572 | гипотетический белок | 95 | 93 | 96 | 9085z | гипотетический белок | 99 | 96 | 100 | |
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31574», «term_id»: «441431490», «term_text»: «AGC31574»} } AGC31574 | 9085 7 портальный белок 97 | 97 | 99 | |||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31576», «term_id» : «441431492», «term_text»: «AGC31576»}} AGC31576 | N-ацетилмурамидаза | 95 | 97 | 98 | IPR008565; IPR018537 | |||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31502», «term_id»: «441431418», «term_text»: «AGC31502»}} AGC31502 | терминазная субъединица A | 98 | 98 | 100 | IPR004921 | |||||
| {«type»: «entrez-protein», «attrs»: {«text3:» AGC3150 term_id «:» 441431419 «,» term_text «:» AGC31503 «}} AGC31503 | гипотетический белок | 98 | 98 | 100 | гипотетический белок | 80 | 91 | 85 | ||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31505», «term_id»: «441431421», «term_text»: «AGC31505»}} AGC31505 | ч yпотетический белок | 81 | 76 | 77 | ||||||
| {«type»: «entrez-protein», «attrs»: {«text»: «AGC31506», «term_id»: «441431422», «term_text»: «AGC31506»}} AGC31506 | гипотетический белок | 87 | 74 | 80 | ||||||
Таблица 2
Общие характеристики и номера доступа Nio4ges к геному бактерий участвуют в мультилокусном филогенетическом исследовании
| Фаг | Источник изоляции | Хозяин | Регистрационный номер | 908| материал | E.coli | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «KC206276», «term_id»: «531936789», «term_text»: «KC206276»}} KC206276 | Это исследование | | |
|---|---|---|---|---|---|---|---|---|---|
| KBNP21 | Куриная ферма | E. coli | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «JX415535», «term_id» «:» 404056358 «,» term_text «:» JX415535 «}} JX415535 | [24] | |||||
| vB_EcoP_G7C | Фекалии лошади | E.coli | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «HQ259105», «term_id»: «343082194», «term_text»: «HQ259105»}} HQ259105 | [25] | |||||
| IME11 | Больничные сточные воды | E. coli | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «JX880034″, » term_id «:» 40 | 00 «,» term_text «:» JX880034 «}} JX880034 | [6] | ||||
| N4 | Канализация | E.coli | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «EF056008», «term_id»: «117606835», «term_text»: «EF056008»}} EF056008 | [11] | |||||
| EamP-S6 | Среда для производства фруктов | Erwinia amylovora | {«type»: «entrez-nucleotide», «attrs»: {«text»: «NC_0_0 , «term_id»: «422935839», «term_text»: «NC_019514»}} NC_019514 | [17] | |||||
| DSS3P2 | Образец морской воды | Silicib : «entrez-нуклеотид», «attrs»: {«text»: «Nc_012697», «term_id»: «237651540», «term_text»: «NC_012697»}} Nc_012697 | [7] | ||||||
| Образец морской воды | Sulfitobacter sp. | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «NC_012696», «term_id»: «237651460», «term_text»: «NC_012696»}} NC_012696 | [ 7] | ||||||
| LIT1 | Пруд | Pseudomonas aeruginosa | {«type»: «entrez-нуклеотид», «attrs»: {«text»: «NC_013_692», «282598847», «term_text»: «NC_013692»}} NC_013692 | [9] | |||||
| LUZ7 | Образец больничных отходов | Pseudomonas aeruginosa | «nucleoti» {тип Pseudomonas aeruginosa: «nucleoti» { «attrs»: {«text»: «NC_013691», «term_id»: «282599392», «term_text»: «NC_013691»}} NC_013691 | [9] |
Выяснение эволюционной взаимосвязи различных N4 -подобные бактериофаги на основе мультилокусных и моногенных филогенетических анализов. Всего для мультилокусного филогенетического анализа использовали семь ортологичных групп. Четыре из семи ортологичных групп содержали функциональное назначение и впоследствии были выбраны для построения индивидуального филогенетического дерева с одним геном.
Выводы
Мы впервые сообщили о последовательности генома N4-подобного бактериофага, который способен инфицировать птичьего патогенного микроорганизма Escherichia coli O78: K80. Анализ его генома выявил новую геномную область, которая может отвечать за специфичность его хозяина.Это исследование также идентифицировало ортологические группы белков, общих для всех секвенированных N4-подобных фагов, и показало, что эволюционное родство N4-подобных бактериофагов коррелирует с характером специфичности хозяина.
Материал и методы
Секвенирование генома и аннотация бактериофага EC1-UPM
Методы размножения, очистки и выделения геномной ДНК бактериофага EC1-UPM соответствуют методам, описанным Lau et al. [4]. Очищенную ДНК секвенировали на Illumina Hiseq 2000 (считывание парных концов 100 пар оснований).Чтения были собраны de novo с использованием Velvet v1.1.07. Для получения полного генома бактериофага EC1-UPM было выполнено последующее создание каркасов и заполнение промежутков. Кодирующие белки последовательности были предсказаны с использованием комбинированной генной модели из GeneMarkS и Glimmer3 [26]. Предсказанные белки были аннотированы BLAST в соответствии с неизбыточной базой данных NCBI. Дополнительные аннотации были впоследствии выполнены с помощью Blast2Go и InterProscan [27,28]. Возможное присутствие тРНК в геноме определяли с помощью tRNAscan-SE v1.23.
Визуализация генома бактериофага EC1-UPM и сравнение с другими близкородственными геномами бактериофага
Аннотированный полный геном бактериофага EC1-UPM визуализировали с использованием CLC Genomics Workbench 6.0 (CLC Bio, Дания). Чтобы идентифицировать близкородственный бактериофаг, BLASTN выполняли на полном геноме бактериофага EC1-UPM по неизбыточной базе данных NCBI. Генератор изображений Blast Ring [29] был использован для обеспечения геномного сравнения бактериофага EC1-UPM с его близкородственным бактериофагом (отсечение 40% идентичности).
In-silico анализ белков хвостовой иглы и хвостовых волокон бактериофага EC1-UPMДополнительная аннотация белка хвостовых волокон была выполнена с использованием интерактивного сервера HHpred [30]. Кроме того, расчет различных физических и химических параметров белка хвостового волокна был выполнен с помощью ProtParam [31].
Классификация холина бактериофага EC1-UPM
Белковая последовательность предполагаемого холина была подвергнута предсказанию трансмембранных спиралей с использованием TMHMM Server v.2.0. Вывод текста прогноза был впоследствии передан в TMRPres2D [32] для создания однородных, двумерных графических изображений высокого анализа предполагаемого трансмембранного белка.
Эволюционное родство N4-подобных бактериофагов
Доступные в настоящее время геномы N4-подобных бактериофагов были получены из NCBI в формате Genbank. PanOCT [33] был использован для идентификации генов, которые являются консервативными в N4-подобных бактериофагах (отсечение идентичности 40% или 70%, перекрытие по длине не менее 70%).Каждый кластер ортологов выравнивали с помощью MUSCLE [34]. Затем индивидуальное выравнивание было обрезано с помощью TrimAl с использованием параметра «strictplus», который оптимизирован для построения дерева NeighborJoining [35] и объединен для филогенетического анализа с использованием MEGA. Филогенетическое дерево было построено с использованием метода NeighborJoining с 1000 повторениями начальной загрузки.
Вклад авторов
Компания HMG провела генетический анализ, подготовила рукопись. CCS разработала весь процесс и помогла редактировать рукопись.SGHT участвовал в экспериментах. ARO и YWH предоставили оборудование и проверили рукопись. Все авторы прочитали и одобрили окончательную рукопись.
Благодарности
Эта работа была поддержана грантом Института биологических наук, Центр передового опыта высших учебных заведений (IBS HICoE) Министерства высшего образования правительства Малайзии. Мы хотим поблагодарить Дж. Л. Лау за ее руководство при подготовке геномного материала и Ю. Л. Тэя (Bioeasy SB) за помощь в области биоинформатики в построении многолокационного филогенетического дерева.
Оценка шести процедур дезактивации для выделения комплекса Mycobacterium avium из фекалий птиц
Ссылка
Менгал, Абдул Саттар и Закария, Зунита и Абу, Джалила и Абдул Азиз, Салеха и Габриэль, Рохас Понсе (2018) Оценка шести процедур обеззараживания для выделения комплекса Mycobacterium avium из птичьих фекалий. PLoS One, 13 (8). С. 1-16. ISSN 1932-6203Аннотация
Культура считается золотым стандартом для окончательной диагностики микобактериальных инфекций.Однако единого мнения о наиболее подходящей методике культивирования для выделения нетуберкулезных микобактерий нет. В исследовании сравнивалось восстановление микобактерий после обеззараживания свежих и колючих фекалий птиц 4% гидроксидом натрия (NaOH), 12% серной кислотой (h3SO4) или 1% хлоридом цетилперидиния (CPC) со смесью трех антибиотиков и без нее, а именно: ванкомицин (VAN, 100 мкг / мл), налидиксовая кислота (NAL, 100 мкг / мл) и амфотерицин B (AMB, 100 мкг / мл). Смесь антибиотиков была обозначена как VNA.Процедуры обеззараживания оценивали с использованием двух (n = 2) образцов фекалий птиц, содержащих 106, 104 и 102 КОЕ / мл Mycobacterium avium subsp. avium (ATCC 15769) и свежие птичьи фекалии (n = 42). М. avium subsp. avium был обнаружен на питательной среде из образцов с добавками (106 и 104 КОЕ / мл), обеззараженных NaOH, NaOH-VNA, h3SO4 и h3SO4 -VNA в течение 2-6 недель. Эти бактерии выявлялись через 2–4 недели при использовании CPC и CPC-VNA. М. avium subsp. avium нельзя изолировать на питательной среде из образцов с добавками (102 КОЕ / мл), деконтаминированных каким-либо обеззараживающим агентом.Два микобактериальных изолята, а именно Mycobacterium terrae и M. engbaekii, были выделены из полевых образцов, деконтаминированных NaOH и CPC-VNA. Что касается скорости загрязнения, использование CPC-VNA показало более низкие уровни загрязнения (5,5% и 19,0%) от проб с добавлением и полевых проб, чем при использовании других методов (NaOH: 22,2% и 59,5%, NaOH-VNA: 16,7% и 21,4%, h3SO4: 11,1% и 40,5%, h3SO4-VNA: 5,5% и 21,4%, и CPC: 66,7% и 50%). В заключение следует отметить, что обеззараживание образцов фекалий в соответствии с двухэтапной процедурой с использованием 1% CPC и VNA может обеспечить высокую скорость восстановления многих микобактерий с минимальным загрязнением культур.
Загрузить файл
Дополнительные метаданные
Действия (требуется логин)
| Просмотреть товар |
Для оценки клинических, биологических и компьютерных критериев для прогнозирования результатов лечения неэкстренной спаечной непроходимости тонкой кишки
- Название:
- Определение клинических, биологических и предиктивных сканографических исследований по проблеме медицинских методов в кадрах окклюзионных дижестивов без первоначальных знаков гравитации
- Автор:
- РУЙЕР, Албан;
- Ответственный за диссертацию:
- CENAC-MILLET, Ингрид;
- Университет:
- Faculté de médecine, Монпелье;
- Год публикации:
- 2013
- Язык:
- французский; Английский ;
- Аннотация:
- Методы: Мы ретроспективно рассмотрели 221 компьютерную томографию пациентов с диагнозом спаечной непроходимости тонкой кишки (ASBO).Были изучены следующие клинические и биологические данные: возраст, пол, предыдущий анамнез висцерального хирургического вмешательства, предыдущий анамнез ASBO, задержка между появлением симптомов и реализацией КТ, задержка между КТ и возможным хирургическим лечением, температура, уровень С-реактивного белка, лейкоцитарный анализ крови и уровень лактатдегидрогеназы. В нашем исследовании оценивались следующие характеристики КТ: наличие назогастрального зонда, расположение обструкции, максимальное растяжение кишечника, соотношение между максимальным диаметром кишечника и дистальным отделом тонкой кишки, прилегающим к переходной зоне, наличие или отсутствие растяжения желудка или проксимального отдела тощей кишки, переднее наложение, наличие признака тонкой кишки и ее длина, наличие клюва и количества клювов, наличие замкнутой петли, С- или У-образных петель, наличие признака завихрения, пристеночного утолщения, наличие недостаточного опорожнения кишечника стенка, брыжеечная инфильтрация и перионеальная инфильтрация.Все эти переменные сравнивались между пациентами, которым было успешно проведено медикаментозное лечение, и теми, кто нуждался в хирургическом вмешательстве. Все переменные, которые повысили значение p <0,2 в одномерном анализе, были введены в многомерную модель, чтобы оценить их независимость. Статистический анализ проводился с использованием компьютерного программного обеспечения (SAS, версия 9.3, SAS Institute Cary, NC). Результаты: на основе многомерного анализа мы показали, что четыре параметра значимо связаны с успехом лечения: продолжительность симптома менее 18 часов (p = 0,003), количество лейкоцитов 8600x106 / л или более (p = O, OI), передняя аппозиция (p = O, OI) и количество клювов (p = 0,005).Заключение Мы показали, что многие выводы могут повлиять на управление неаварийной ASBO. Было показано, что пациенты с числом лейкоцитов выше 8600х106 / л, с числом клювов меньше или равным 1 и продолжительностью симптомов менее 18 часов, успешно прошли курс лечения. Мы обнаружили новый признак: переднее положение, которое ранее не описывалось, и оно было очень тесно связано с успехом безоперационного лечения. ;
- Аннотация:
- Методы: Nous avons relu rétrospectivement 221 сканер пациентов принимает дополнительные меры! une occlusion aigue du grêle sur phénomènes adhérentielles.Les Critères Scanographiques Notés Furent: présence d’une sonde naso-gastrique, localization de l’occlusion, растяжка максимального du grêle, rapport entre le diamètre maximal du grele et l’anse d’aval à la zone de transition, présence ou non d ‘une extension du gastrique ou jéjunaleximale, apposition antérieure. Все эти переменные сравниваются с пациентами, которые достигли успеха: в медицине и в других странах, где проводится лечение без хирургических вмешательств. Результаты: Basé sur l’analyses multitivariée, nous avons montré que quatre paramètres étaient Mean Signative associés à la réussite du traitement médical: la durée des симптомы de moins de 18 heures (p = 0,00×103), le taux plus de leucocyte 8600 (p = 0,01), un apposition antérieure (p = 0.01) et le nombre debes (p = 0,005). Заключение Nous avons Trouvé un nouveau signe: l’apposition antérieure, qui n’a jamais été décrit auparavan et il été associé très fortement à un succès du traitement médical. ;
- Пагинация / Размер:
- 1 т., 44 с., Библиогр. f. 41-44 (72 см.), Иллюстрация;
- Классификация SIGLE:
- 060 — Биологические и медицинские науки, общие;
- Ключевое слово:
- Медицина; Médecine; Сканография; Imagerie par résonance magnétique; Imagerie для диагностики; Intestin grêle; Кишечная непроходимость; Диссертации университарии; Apposition antérieure;
- Дисциплина:
- Médecine
- Тип документа:
- У — Диссертация; (Тезис)
- Отношение:
- http: // www.sudoc.fr/171435966
- Другой идентификатор:
- , номер диссертации: 13МОН11014; inist: 27729792;
- ручка:
- http://hdl.handle.net/10068/894827
- Происхождение:
- Sudoc;
- Получить копию:
- Судок
- Наличие:
- MONTPELLIER-BU Médecine UPM (341722108)
- Страна:
- Франция;
Присутствие живого вируса SARS-CoV-2 в кале от коронавирусной болезни 2019 (COVID-19) Пациенты: быстрый обзор
РЕЗЮМЕ
Коронавирусная болезнь 2019 (COVID-19) — это быстро развивающаяся пандемия, которая распространилась на многих части света.Болезнь уже затронула более 6 миллионов человек, более 400 000 человек погибли. Недавние исследования подтвердили наличие нуклеиновых кислот SARS-CoV-2 в кале пациентов с коронавирусной болезнью 2019 (COVID-19) с помощью тестов RT-PCR. Однако до сих пор неясно, присутствует ли живой вирус SARS-CoV-2 в фекалиях этих пациентов. В этом быстром обзоре мы систематически проанализировали литературу, чтобы установить любые доказательства наличия живого вируса SARS-CoV-2 в образцах фекалий пациентов с COVID-19.Мы выявили 4 исследования (одно клиническое наблюдение, 2 серии случаев и 1 когортное исследование), в которых SARS-CoV-2 был успешно выделен из образцов фекалий пациентов с COVID-19 с использованием методов культивирования. Таким образом, есть некоторые свидетельства того, что COVID-19 может распространять живой вирус SARS-CoV-2 через желудочно-кишечный тракт. Необходимы более масштабные исследования, чтобы подтвердить эти выводы, а также определить его потенциал для передачи заболеваний и инфицирования, а также возможные последствия для политики выделения и изоляции COVID-19.
Заявление о конкурирующем интересе
Авторы заявили об отсутствии конкурирующего интереса.
Отчет о финансировании
Автор (ы) не получил финансовой поддержки для исследования, авторства и / или публикация этой статьи.
Заявления авторов
Я подтверждаю, что были соблюдены все соответствующие этические принципы и получены все необходимые разрешения IRB и / или комитета по этике.
Да
Подробная информация об IRB / надзорном органе, предоставившем разрешение или исключение для описанного исследования, приведена ниже:
Не применимо
Получено все необходимое согласие пациента / участника, а соответствующие институциональные формы заархивированы.
Да
Я понимаю, что все клинические испытания и любые другие проспективные интервенционные исследования должны быть зарегистрированы в одобренном ICMJE реестре, таком как ClinicalTrials.gov. Я подтверждаю, что любое такое исследование, указанное в рукописи, было зарегистрировано и предоставлен идентификатор регистрации исследования (примечание: при публикации проспективного исследования, зарегистрированного ретроспективно, просьба предоставить заявление в поле идентификатора исследования, объясняющее, почему исследование не было зарегистрировано заранее) .
Да
Я выполнил все соответствующие инструкции по составлению отчетов об исследованиях и загрузил соответствующие контрольные списки отчетов по исследованиям сети EQUATOR и другие соответствующие материалы в качестве дополнительных файлов, если применимо.
Да
Сноски
Заявление о конфликте интересов: Ни у одного из авторов нет конфликта интересов в отношении данной публикации.
Финансирование: Автор (ы) не получил финансовой поддержки для исследования, авторства и / или публикации этой статьи.
Доступность данных
Все подтверждающие данные находятся в рукописи
Когда кал — лучшее лекарство
В течение двух-трех дней после трансплантации у большинства пациентов «симптомы отсутствуют … Они возвращаются к своей жизни», — сказал Майкл. Эдмонд, специалист по инфекционным заболеваниям в Университете Содружества Вирджинии.Это почти чудодейственное лекарство, которое предлагает медицина.
Тем не менее, доступ к трансплантации фекалий оказался проблематичным. Еще в 2013 году Эми Барто, гастроэнтеролог из клиники Лахи в Массачусетсе, сказала, что ее пациенты должны были найти собственных доноров стула, которых клиника будет проверять индивидуально. В день процедуры донор должен был предоставить свежий образец стула, который, по словам Барто, она лично смешала с помощью блендера от Target и трансплантировала в толстую кишку пациента. «Пациентам было неловко и нервно искать собственных доноров, а обследование — дорогое удовольствие», — сказала она.«Я проделал около 100 процедур с помощью блендера, и это было неэффективно».
В 2013 году Смит и его друг по колледжу Джеймс Берджесс решили открыть некоммерческий банк стула под названием OpenBiome, предоставляющий предварительно проверенные замороженные образцы врачам и больницам. Консультант Смита из Массачусетского технологического института Эрик Алм предложил руководство, а также уголок его лаборатории, и команда начала набор доноров, в основном из числа исследователей Массачусетского технологического института и Гарварда. «Когда был создан OpenBiome, качество моей жизни резко возросло», — сказал Барто.Что еще более важно, доступ к процедуре «просто расцвел». Больше врачей были готовы принять участие, и пациенты смогли пройти процедуру быстрее, с меньшими препятствиями и меньшими затратами.
Одной из таких пациенток была Натали Джамиль. В апреле 25-летняя Джамиль обнаружила, что корневой канал, который у нее был в течение многих лет, сильно инфицирован. Дантист удалил зуб, назначив ей антибиотики до и после процедуры. Однако через неделю после того, как она прекратила прием препарата, у нее началась рвота, возникла сильная диарея, спазмы и боли в суставах.Джамиль недавно устроилась на работу секретарем в Вашингтоне, округ Колумбия, но «работать стало невозможно, и меня попросили уйти в отставку», — сказала она. «Я остался без работы, без медицинской страховки и без зубов».
В отчаянии поискав в Интернете, она нашла Эдмонда. «Я потерял более 25 фунтов за два месяца», — сказал Джамиль. «Я был готов на все».
В июне Джамиль прошел курс лечения замороженным стулом от OpenBiome. Эдмонд ввел образец через трубку, которая прошла через нос и попала в тонкий кишечник.«Это было похоже на мерцание света», — сказала она. «На следующий день я снова встал на ноги. Мой желудок был спокойным. У меня не было диареи. Это было похоже на волшебство ».
* * *
OpenBiome в некотором смысле похож на банк крови, который также собирает, проверяет и распределяет биологический материал. Однако на раннем этапе стало ясно, что группа сталкивается с разными проблемами при наборе доноров и тестировании образцов. С одной стороны, подходили немногие кандидаты: любой, кто болел желудочно-кишечным заболеванием, недавно принимал антибиотики или путешествовал в любую страну с риском заболеваний, передающихся через воду, был отклонен еще до того, как их стул был проанализирован на патогены.Иногда было неловко отвергать коллег из Массачусетского технологического института, которые хотели внести свой вклад, сказал Смит, но «это помогло сказать:« Эй, я тоже не имею права »» (брат Смита живет в Сингапуре, и он приезжает туда каждый год). OpenBiome проверяет стул на наличие обширного списка патогенов, включая ВИЧ, гепатит и сифилис, которые могут передаваться через физиологические жидкости. Но они также выявляют расстройства, от ожирения до метаболического синдрома и аутоиммунных заболеваний, которые традиционно не считаются заразными, но, по крайней мере, коррелируют с нарушениями кишечных бактерий.В банке крови никто не беспокоится о том, что переливание крови может вызвать ожирение.
Препараты фекальной микробиоты OpenBiome на складе(Кэролайн Эдельштейн)
Вместо того, чтобы расширять свою сеть для доноров, OpenBiome культивирует небольшую группу стойких участников. Таким образом, стоимость тестирования, которая может составлять примерно 1500 долларов на донора, распределяется между многими пациентами и видами лечения. Участники этой модели берут на себя серьезные обязательства. От них требуется сдать образцы в течение часа после прохождения, поскольку бактериальный состав стула быстро меняется при контакте с воздухом.